
 Los hallazgos de 19 genomas humanos antiguos de la región del lago Baikal, incluido este niño de Mal´ta, uno de los más antiguos con 24.000 años de la región, y que dan un poco de luz sobre las migraciones antiguas. Mientras que algunos individuos tenían claramente un genotipo del este de Asia, el genoma del chico de Mal'ta es el humano moderno más antiguo de aquella zona y muestra que 1/3 de su ascendencia es europea, presentándose con una cultura material nada común en Asia.
Los hallazgos de 19 genomas humanos antiguos de la región del lago Baikal, incluido este niño de Mal´ta, uno de los más antiguos con 24.000 años de la región, y que dan un poco de luz sobre las migraciones antiguas. Mientras que algunos individuos tenían claramente un genotipo del este de Asia, el genoma del chico de Mal'ta es el humano moderno más antiguo de aquella zona y muestra que 1/3 de su ascendencia es europea, presentándose con una cultura material nada común en Asia.
El sitio de Mal'ta está ubicado en la orilla izquierda del Belaya, un afluente del Angara, a su vez un afluente del Yenisei, y está a cien kilómetros al noroeste de Irkutsk y el lago Baikal. En los alrededores del lago existen numerosas fuentes de aguas termales, como las de Arshán o Goryachinsk, que son populares balnearios para el turismo de salud. Y se han detectado emanaciones de calor y fluidos hidrotermales en el fondo del lago, lo que contribuye a que sus aguas profundas no se estanquen y mantengan niveles altos de oxígeno.
Descubierto en 1928, fue objeto de numerosas excavaciones sucesivas por Sergei N. Zamiatnine, GP Sosnovskii y, especialmente, por el señor Mijail Gerasimov, que trabajó allí durante más de treinta años. Aunque la ocupación humana más temprana de Mal´ta probablemente comenzó hace unos 40.000 años. Pequeños grupos de cazadores de caza mayor probablemente migraron a esta región desde tierras al sur y suroeste, enfrentándose a un clima duro e inviernos largos y secos. Parece que Mal'ta es el yacimiento más antiguo conocido en Siberia oriental, junto con el cercano yacimiento de Buret'.
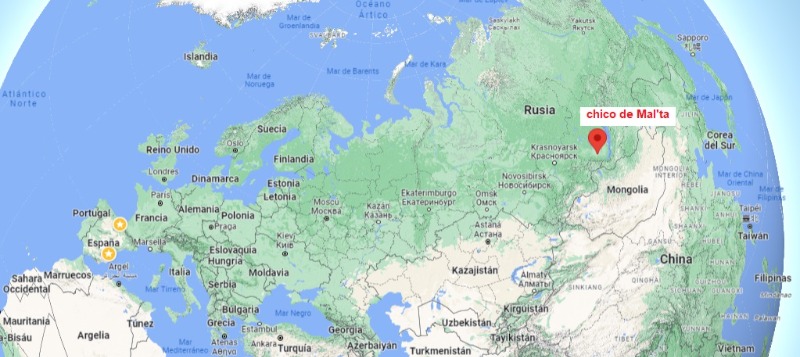
Un niño, cuyos restos fueron hallados enterrados bajo el suelo de una vivienda de Mal'ta, se le conoce generalmente por la abreviatura MA-1, y los restos han sido datados en 24 000 años antes del presente. El entierro del niño en Mal'ta es de particular interés al considerar la evidencia de la infancia en el registro paleolítico de Siberia. El esqueleto fue enterrado dentro de una estructura de losa que se asemeja a una tumba excavada y revestida. El fondo del entierro estaba cubierto de ocre rojo. Se encontraron varios adornos y objetos utilitarios entre los huesos: fragmentos de una tiara hecha de marfil de mamut, un rico collar de 120 cuentas de hueso, un gran colgante ovalado, una placa de hueso redonda con un patrón en zig-zag y una escultura de un pájaro volando. Además de los joyeros, varias herramientas de piedra, así como un cuchillo de sílex toscamente procesado, yacían junto al niño. Gerasimov estaba firmemente convencido de que este inventario de artefactos no podía pertenecer al propio niño debido a su edad, de 3 a 4 años.

MA-1 es el ÚNICO ejemplo conocido de ADN-Y “basal” R, es decir, el único miembro del haplogrupo paterno R que NO PERTENECÍA a los haplogrupos R1, R2, ni a subclados secundarios de estos. Su ADN mitocondrial (materno) pertenecía a un subclado no resuelto del haplogrupo U.
Según investigaciones publicadas desde 2013, MA-1 pertenecía a la población de los antiguos euroasiáticos del norte, quienes eran genéticamente «intermedios entre los euroasiáticos occidentales modernos y los nativos americanos, pero distantes de los asiáticos orientales», y parcialmente "ancestro genético de los siberianos", los indígenas americanos, los pueblos yamnaya y botai de la Edad del Bronce de la estepa euroasiática.
Reich habla de una población que aportó genes tanto a los amerindios como a los euroasiáticos occidentales. Reich denomina a esta población como "asiática", pero bien podría llamarse de otra forma, ya que no hay nada en los datos que nos indique que esta población ancestral estuviera en Asia, ya que no existe relación entre Mal'ta y los asiáticos orientales modernos.
¿Es la de Mal´ta una gente originaria de la estepa siberiana?
Gran parte de lo que se sabe sobre Malta proviene del arqueólogo ruso Mijaíl Gerasimov. Gerasimov realizó descubrimientos revolucionarios al excavar Malta en 1927. Hasta entonces, las sociedades del Paleolítico Superior del norte de Asia eran prácticamente desconocidas. El arte mueble, generalmente algún tipo de talla en marfil, colmillo o asta, se extendía desde Europa Occidental hasta el norte y centro de Asia.

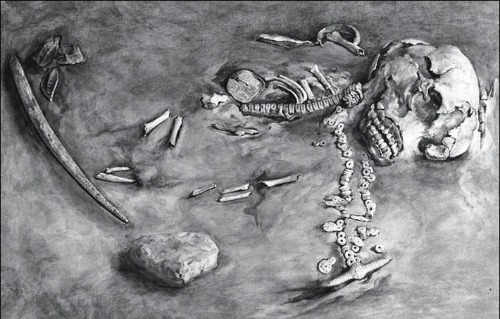
Los restos artísticos de objetos de hueso, marfil y asta, tallados con gran maestría y que representan aves (cisnes) y figuras femeninas, son los más comunes; estos objetos constituyen, en conjunto, la principal fuente de la fama de Mal'ta. Contaban también con fragmentos de jade, cuchillos hechos con hojas, puntas de hueso y finas figuras de marfil: esculpidos colimbos y cisnes. En las casas se encontraron collares de marfil de mamut, colgantes de calcita, botones decorados con líneas en zigzag talladas, agujas, punzones, raspadores y cuchillos.

En un colmillo de mamut se muestra la figura de un mamut, fácilmente reconocible por su trompa, colmillos y patas gruesas. La lana también parece estar grabada, a juzgar por las líneas rectas que recorren el cuerpo. Otro dibujo representa tres SERPIENTES con la cabeza erguida y ladeada. En un lado de un botón o colgante de placa podemos ver tres serpientes. La serpiente es rara en el arte paleolítico del hemisferio norte, presumiblemente porque las condiciones de frío impidieron una amplia distribución de serpientes.
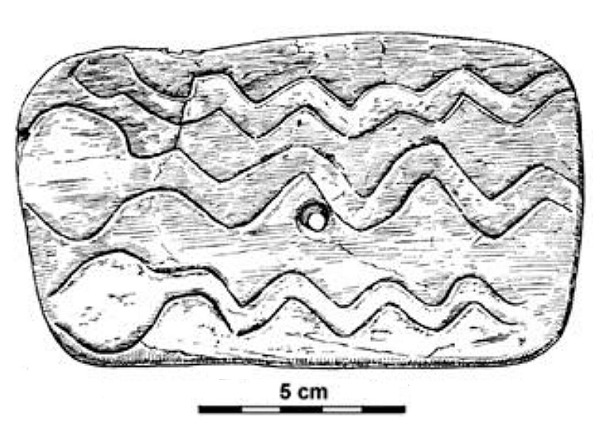
De vez en cuando había una figura femenina, que suscitan interés en Occidente por su aparente SIMILITUD MORFOLÓGICA CON LAS FIGURILLAS FEMENINAS EUROPEAS DE LA MISMA ÉPOCA, lo que sugiere una posible conexión cultural y de culto. Esta similitud entre Mal'ta y el Paleolítico Superior Europeo coincide con otras semejanzas observadas entre ambas culturas, como sus herramientas, el uso de pigmento rojo (ocre o cinabrio)
Sin embargo, carece de los típicos skreblos (grandes raspadores laterales) que son comunes en otros yacimientos paleolíticos siberianos. Además, nunca se han encontrado otras características comunes, como núcleos de guijarros, núcleos en forma de cuña, buriles y herramientas compuestas. La falta de estas características, combinada con un estilo artístico que solo se encuentra en otro yacimiento cercano, hace que la cultura Mal'ta sea RARA Y ÚNICA en Siberia.
Mal'ta consiste en viviendas semisubterráneas construidas con huesos de mamut, huesos de animales grandes para las paredes y astas de reno recubiertas de pieles para el techo. Construcciones similares a las encontradas en el Paleolítico Superior de Eurasia occidental, como en zonas de Francia, Checoslovaquia y Ucrania.
Estudios genéticos
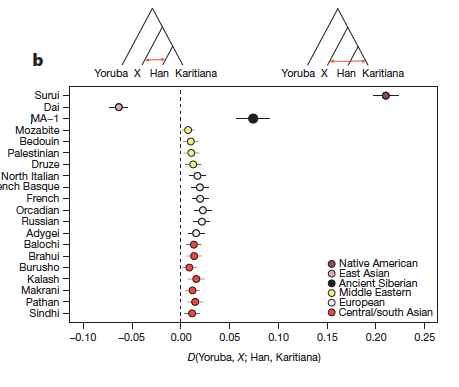
Los resultados de estudios genéticos de estos restos humanos de Siberia muestran que, personas emparentadas con europeos occidentales se extendieron más al este de lo sospechado y vivieron en Siberia durante las partes más frías de la última Edad de Hielo. Y así, algunos asiáticos orientales y europeos occidentales dieron lugar a una mezcla genética, que según el paleontólogo Eske Willerslev, creó a los nativos americanos, siendo los predecesores de las poblaciones que posteriormente llegaron a América, tanto al norte como al sur. En la siguiente imagen vemos los grupos poblacionales más cercanos genéticamente a MA-1, entre ellos Escoceses de las Orcadas, rusos, franceses y vascos.
DNA materno.
En 2009, el equipo de Willerslev recogió una muestra de ADN del chico de Mal'ta. El equipo encontró que el ADN (de 24.000 años) y las mitocondrias del joven pertenecían a un linaje materno de haplogrupo U, que, si bien se encuentra en Europa y en el oeste de Asia, no es natural verlo en el este de Asia donde fue descubierto este cuerpo. El haplogrupo U también se ha encontrado con alta frecuencia entre los cazadores-recolectores europeos del Paleolítico superior y el Mesolítico.
El mapa a continuación, del material complementario incluido en el artículo, muestra la distribución del haplogrupo U; los puntos negros indican las ubicaciones del ADN de comparación del haplogrupo U.
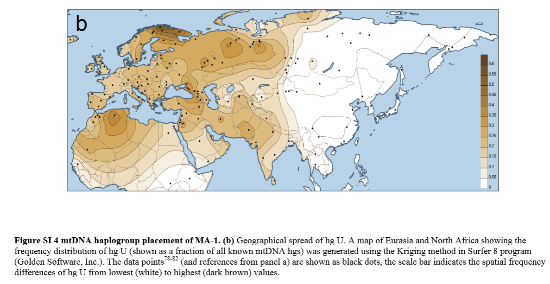
Con haplogrupo U encontraron en Bélgica los restos de una mujer (GoyetQ53-1) de 28.500 años y en la República Checa otra mujer de 31.000 años. La amplia distribución geográfica del haplogrupo U en el Paleolítico Superior y Mesolítico va desde sitios en Portugal (Chandler et al., 2005), y los dos individuos de LaBraña en León en el Oeste Atlántico, hasta el sitio de Yuzhny Oleni Ostrov en el norte de Karelia (Der Sarkissian et al., 2013), Kostenki en Rusia (Krause et al., 2010) y el niño Mal'ta en la región del lago Baikal (Raghavan et al., 2014) indica un tamaño de población relativamente pequeño de grupos de cazadores-recolectores, que se extendían a lo largo de un vasto espacio longitudinal de Europa y Asia. En un artículo de Brandt et al., publicado en octubre de 2013, los autores informan que, en las 198 muestras de ADN antiguo recolectadas en 25 yacimientos alemanes de cazadores-recolectores antiguos correspondieron a los haplogrupos U, U4, U5 y U8. No se encontró representación de ningún otro haplogrupo.
El genoma del individuo del yacimiento La Braña , en España (el más antiguo de la prehistoria recuperado en España) y según el investigador Carles Lalueza-Fox, mostró que La Braña 1 "tiene un ancestro común con los pobladores del yacimiento del Paleolítico superior Mal'ta, ubicado en el lago Baikal (Siberia)” y "posiblemente están estrechamente relacionados con Mal'ta" según Sánchez-Quinto.
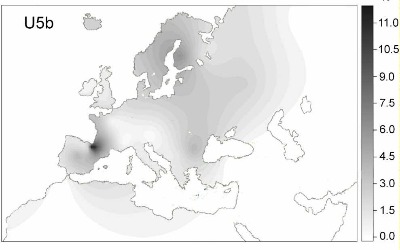
Frecuencias del Haplogrupo U5 en Europa según Eupedia:
Los Sami (47,5%).
Norte de Navarra (15,5%)
Noruega, Suecia y Dinamarca (6% y 15 %)
Letones en el Báltico (13,5%)
Estonios (13%)
Ávaros (13%)
Vascos (12%)
Lituanos (11,5%)
Cántabros (11%)
Eslovacos (11 %)
Bretones (10,5 %)
Tártaros, Rusia (10,5%)
Croatas (10,5 %)
Chechenos, catalanes, sardos, polacos, checos, ucranianos, rusos eslavos (10%)
Marruecos (4 %)
Libia (3,5 %)
Cáucaso (3,5 %)
Irán, Turquía (3 %)
Túnez y Argelia (ambos con un 2 %)
Siria, Líbano, Irak y Egipto (alrededor del 1,5 %)
Si algún portador del haplogrupo U afirma tener un parentesco lejano con MA-1, quien vivió hace 24.000 años y que también estaba emparentado con su ancestro común, quien vivió en algún momento anterior, entre 24.000y 50.000 años atrás, probablemente en algún lugar entre Oriente Medio, donde nació U, Malta, Siberia y Europa Occidental, estaría en lo cierto.
DNA paterno.
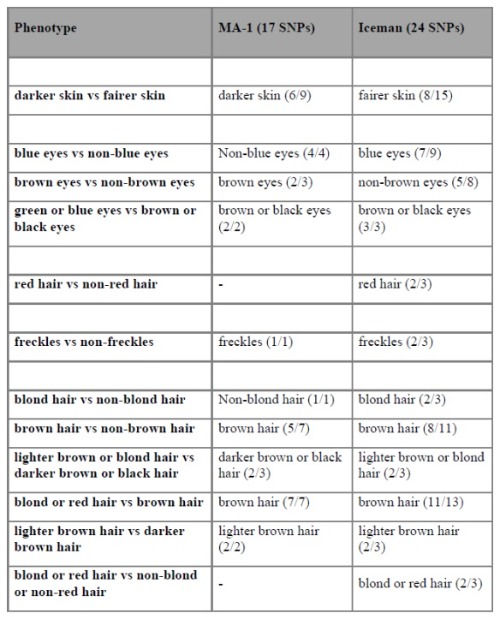
El ADN del chico demostró que genéticamente, esta persona NO TENÍA NINGUNA SEMEJANZA CON LASPOBLACIONES DE ASIA ORIENTAL, pero se parecía a los europeos que se habían trasladado a Asia occidental, destacó Willerslev. Los autores compararon la información del fenotipo obtenible de MA-1 con el Hombre de Hielo, típicamente llamado Otzi. Esto es particularmente interesante a la luz del cambio de pigmentación de piel más oscura en África a piel más clara en Eurasia, y la pregunta de cuándo ocurrió este cambio de apariencia. MA-1 muestra una afinidad genética con las personas contemporáneas del norte de Europa, la población hoy en día con la frecuencia más alta de fenotipos de pigmentación clara.
Los autores compararon el ADN de MA-1 con un conjunto de 124 SNP identificados en 2001 por Cerquira como informativos sobre el color de la pigmentación de la piel, el cabello y los ojos, aunque también advierten que este método tiene una precisión de predicción limitada. Dado eso, dicen que MA-1 tenía cabello, piel y ojos oscuros, pero no pudieron secuenciar el conjunto completo de SNP. MA-1 también presentó el valor de SNP asociado con un alto riesgo de calvicie de patrón masculino y no presentó tolerancia a la lactosa, un rasgo presente en los euroasiáticos occidentales.
MA-1 tampoco presenta la mutación asociada con el grosor del cabello y los incisivos en forma de pala en los asiáticos.
El niño Mal'ta tenía además un haplogrupo (paterno) R basal , y probablemente es anterior a la división en los subgrupos R1a, R1b y R2. Es el Y cromosoma tipo R más antiguo localizado hasta el momento, de las “solo “cuatro antiguas aparecidas.
El genoma del Niño de Mal'ta también mostró que los nativos americanos pueden remontar entre un 14/38 % de su ascendencia del oeste de Europa. El equipo de Willerslev sugiere que después de que los antepasados de los nativos americanos se separasen de los de los asiáticos orientales, se trasladaron hacia el norte. En algún lugar de Siberia, se encontraron con otro grupo de personas que vienen del oeste de Eurasia, población a la que pertenecía el chico de Mal'ta. Los dos grupos se mezclaron, y sus descendientes finalmente viajaron al este en América del Norte. A esa mezcla se llama “ANE” (euro-asiáticos nórdicos antiguos).
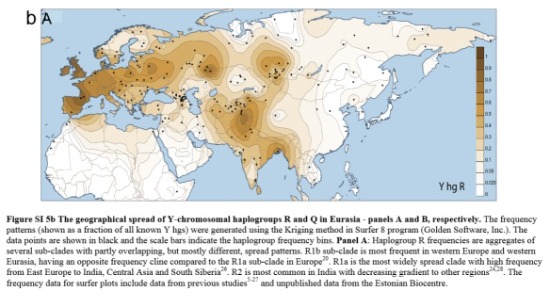
ASCENDENCIA ANTIGUA (ANE)
El término ANE es el nombre dado en arqueo-genética al componente genético ancestral asociado con la cultura de Mal'ta-Buret.
Este componente genético desciende parcialmente de los euro-asiáticos orientales, pero la mayoría de la ancestría genética de los ANE está congeniada con poblaciones euro-asiáticas occidentales como los cazadores-recolectores occidentales (WHG), labradores neolíticos europeos (ENF) y cazadores-recolectores del Cáucaso (CHG, con mayoría de ascendencia basal euroasiática occidental).
"esa presencia oriental en Asia de una población emparentada con los euroasiáticos occidentales contemporáneos ofrece la posibilidad de que las características craneales no orientales de Asia de los primeros americanos se derivaran del Viejo Mundo",, " la presencia de una antigua firma genómica euroasiática occidental en el área del Baikal antes y después del UCM sugiere que partes del centro-sur de Siberia estuvieron ocupadas por humanos durante las etapas más frías de la última edad de hielo"
Roberta Estes , científica, genealogista genética.
R1b es de EUROPA OCCIDENTAL?
El grupo genético R1b es más frecuente en Europa occidental hoy en día y, como no tenemos donantes antiguos, las hipótesis y la fantasía se mezclan en este tema.
La presencia de R1b en Villabruna (Italia) hace 14000 años y en Iberia de 7.000 años documenta una historia más antigua de este haplotipo en el occidente de Europa. El individuo de Villabruna con YDNA era R1b y mtDNA U5b, pertenecía a la cultura gravetiense, cultura que fue reemplazada en iberia y Francia por el solutrense.
No se puede asociar este grupo genético a la llegada de la agricultura, de hecho en los yacimientos AGRÍCOLAS TEMPRANOS no se han encontrado los haplogrupos predominantes europeos modernos, R1b (Europa occidental) y R1a (Europa del este) según Balaresque et al., 2010; Underhill et al., 2010; Myres et al.,2011. En cambio se han encontrado R1b en contextos del Neolítico tardío con una distribución geográfica sorprendentemente coincidente de la cultura arqueológica asociada. Eso significa que los que introdujeron la agricultura en Europa no portaban los tipos R1b o R1a, a la vez, los cazadores recolectores al reconvertirse en agricultores y expandir su población, sí que portaban este genotipo.
Estudios recientes han encontrado que el ADN-Y de los primeros agricultores europeos es típicamente el haplogrupo G2a. [Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA, et al. (Diciembre de 2015). "Patrones de selección de todo el genoma en 230 antiguos euroasiáticos" . Naturaleza. 528 (7583):499–503]
Aun así, se hacen hipótesis de su posible origen y predominio actual del R1b en Europa occidental, mezclándolas con las migraciones de la Edad del Bronce (2,5 / 3 Mil años) desde la estepa de Europa oriental y los Yamnaya. La suposición matemática de multiplicaciones en secuencias del genoma no es la formula es la más adecuada, dando horquillas desde 26.000 a 2.600 años.
Hemos visto que el R del niño de Mal´ta no es un grupo originario de Siberia, estando fuera de lugar incluir su origen estepario. Algunas hipótesis olvidan hoy la rama genética del hombre de Villabruna (mTdna materno U5b2b) con Ydna paterno R1b de hace 14.000 años situada en Italia.
 Hoy sabemos que las muestras antiguas de Yamnaya están ubicadas en la rama "oriental" R-GG400 del haplogrupo R1b-L23, lo que muestra que los descendientes paternos de los Yamnaya todavía viven en la estepa póntica y que la antigua población de Yamnaya no era una fuente importante de linajes paternos en los europeos occidentales de hoy, según BALANOVSKY, O.; CHUKHRYAEVA, M.; ZAPOROZHCHENKO,V. (2017, "La diferenciación genética entre poblaciones de tierras altas y bajas da forma al paisaje cromosómico Y de Asia occidental" (https://labs.icb.ufmg.br/lbem/pdf/Balanovsky2017HGlowlandAsia.pdf )
Hoy sabemos que las muestras antiguas de Yamnaya están ubicadas en la rama "oriental" R-GG400 del haplogrupo R1b-L23, lo que muestra que los descendientes paternos de los Yamnaya todavía viven en la estepa póntica y que la antigua población de Yamnaya no era una fuente importante de linajes paternos en los europeos occidentales de hoy, según BALANOVSKY, O.; CHUKHRYAEVA, M.; ZAPOROZHCHENKO,V. (2017, "La diferenciación genética entre poblaciones de tierras altas y bajas da forma al paisaje cromosómico Y de Asia occidental" (https://labs.icb.ufmg.br/lbem/pdf/Balanovsky2017HGlowlandAsia.pdf )
Origen de R1b.
Un origen propuesto para R1b, y una distribución geográfica actual, es muy similar al del haplogrupo H del ADNmt (Myres et al., 2011). El origen de H se ha atribuido al Cercano Oriente durante el Paleolítico superior, de donde se separó de sus clados hermanos R0 y HV0 al entrar en Europa hace muchos miles de años.
Este haplogrupo H ha sido encontrado con alta frecuencia (38%) en cazadores-recolectores en Portugal y el País Vasco (Chandler et al., 2005; Hervella et al., 2012) y en un individuo del sitio Yuzhnyy Oleni Ostrov en Kareliadel Norte (Der Sarkissian et al., 2013), lo que indica una distribución generalizada y posiblemente clinal del haplogrupo H en la Europa mesolítica.
La mayoría de los subgrupos en la Europa actual muestran fechas de coalescencia glacial tardía o posglacial, lo que aboga por una nueva expansión durante las principales fases de calentamiento después de 15 M. años (Achilli et al., 2004; Pereira et al., 2005; Soares et al., 2010). Y se estima que los subgrupos H1, H3 y H5 se propagaron desde un refugio occidental Franco-Cantábrico basado en fechas en gran medida superpuestas (11,1 kya, 11,5 kya, 13,9 kya, respectivamente). El haplogrupo H de individuos del Mesolítico Ibérico, la resolución de tipificación desafortunadamente no proporciona una asignación inequívoca a ninguno de estos subgrupos (Chandler et al., 2005; Hervella et al., 2012).
Aunque en el estado actual de la investigación, la presencia de los haplogrupos N, H, U5b y U4 (aunque en baja frecuencia) y la ausencia de U2 y U5a, podrían interpretarse como el sustrato genético mesolítico del suroeste de Europa. Las conclusiones hay que sacarlas sin fantasear, pero solo hace falta ver un dato: con la media estimada de 20 km/día de un cazador-recolector, en 365 días llegara al lago Baikal desde los montes pirineos (7.300 km), y en aquella Europa sabemos que se movían mucho.
PESTE
Con ellos también migraban patógenos como la Yersinia pestis, que provoca la peste bubónica y la pulmonar.
Los científicos encontraron restos del bacilo en dos individuos siberianos de la Edad de Bronce Temprano. Las cepas que portaban estos individuos están relacionadas con una cepa contemporánea identificada en un individuo de la región báltica europea, muy al oeste, lo que respalda aún más la alta movilidad de largo alcance tanto de esos patógenos como probablemente de las personas. Aunque se difundió la hipótesis de la propagación de Y Pestis por las migraciones desde la estepa hacia el oeste europeo, los dos individuos aquí identificados con el patógeno eran genéticamente similares al noreste de Asia, pero el análisis de isótopos reveló una señal no local, lo que sugiere los orígenes del patógeno fuera de la región del descubrimiento.