
 El genetista Dr David K. Faux estudio referencias históricas, datos lingüísticos y arqueológicos para vincular los pueblos celtas Hallstatt y LaTene de Europa Central con un marcador de cromosoma Y, el R1b-U152, conectado con el refugio glaciar franco-cantábrico. Para entender la dimensión de este parámetro genético se ha de poner como ejemplo su diversidad, el cantante, compositor y bailarín estadounidense Michael Jackson (el Rey del Pop) perteneció al haplogrupo R1b-U152, al igual que Abraham Lincoln o el actor Kevin Costner.
El genetista Dr David K. Faux estudio referencias históricas, datos lingüísticos y arqueológicos para vincular los pueblos celtas Hallstatt y LaTene de Europa Central con un marcador de cromosoma Y, el R1b-U152, conectado con el refugio glaciar franco-cantábrico. Para entender la dimensión de este parámetro genético se ha de poner como ejemplo su diversidad, el cantante, compositor y bailarín estadounidense Michael Jackson (el Rey del Pop) perteneció al haplogrupo R1b-U152, al igual que Abraham Lincoln o el actor Kevin Costner.
Los pueblos de lenguas indoeuropeas (latinos, germanos, eslavos, celtas, griegos...) se originaron en Europa Occidental, según la "hipótesis bélica" originados por la expansión de los pueblos pastores nómadas yamna (ucranianos, yama significa ‘hoyo’ en ucraniano) en el 2500 a.C.
Otra teoría propone que se dieron en el 4500 a.C. asociado con la expansión de la agricultura en Europa. El asentamiento de humanos portadores de la agricultura (indoeuropeos) en Europa, dio lugar a una expansión demográfica de los indígenas europeos (cazadores y ganaderos) reconvertidos a agricultores estacionales, dándose la inevitable mezcla con los inmigrantes indoeuropeos.
Las lenguas pre-indoeuropeas (posibles hermanos del vasco-ibérico) que pudieron existir en Europa, desaparecieron como tales y otras se fusionaron o mutaron. Actualmente solo quedan un par de idiomas primigenios emparentados con aquella vieja Europa, el fino-ugrio y el euskera.
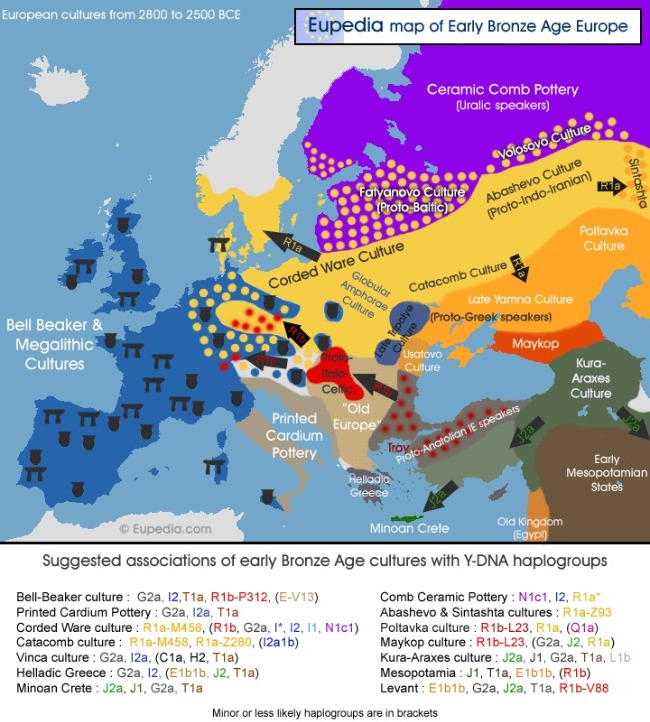
Uno de los primeros pueblos indoeuropeos que se expandieron en Europa fueron los pueblos celtas. Algunos filólogos especialistas en el proto-celta (lengua origen de las actuales lenguas celtas que ha sido reconstruida) defienden la teoría de un sustrato vascónico en esta lengua, dado que encuentran palabras que no tienen origen indoeuropeo y ven en ellas un origen proto-vasco por su cercanía con palabras vascas.
Uno de ellos es el profesor de fonética en la Universidad de Turku, Finlandia, Kaino Kalevi Wiik (Finlandia, 1932-2015). El lingüista finlandés planteó que la cultura megalítica de lengua vasca llegó al área occidental de la cultura Funnel-Beaker (asociado a zona Germánica, Báltica y Eslava), y su resultado fue el desarrollo de un dialecto en una lengua germánica independiente. El dialecto oriental de la cultura Funnel-Beaker se convirtió en un idioma balto-eslavo independiente. El idioma balto-eslavo recibió un sustrato diferente al vascónico, el de los idiomas en la región del Mar Báltico y la región del Mar Negro, y el resultado fue la división del idioma balto-eslavo en dos idiomas, báltico y eslavo. También planteó que los dialectos centroeuropeos recibieron un sustrato vascónico en Europa central, y el resultado fue el celta centroeuropeo (cultura de Michelsburg).
El dialecto mediterráneo y la lengua ítalo-celta, recibió dos tipos de sustrato; En Iberia el resultado fue el celtibero, en Italia el italiano.
El celta centroeuropeo y el celta ibérico formaron una familia de dialectos, que se extendía desde Europa central hasta Iberia, las "lenguas celtas".
¿Quiénes eran los celtas?
Aproximadamente en el 720 al 600 a.C. se dieron en Europa una peculiar forma de enterramiento de la élite celta en los pueblos HalstattC en la región de los Alpes orientales (donde se enriquecieron con el comercio de la sal). Enterramientos al estilo faraónico incorporando a la tumba joyas, armas, carros, mobiliario de hogar, caballos, etc. y dando así una seña de identidad característica a este pueblo. A partir del año 600 a.C., aproximadamente, la base de poder se desplazó hacia el oeste, a las áreas de HallstattD en el sur de Alemania, Suiza y el este de Francia. Hacia el 480 a.C. estos entierros de élite reflejan el establecimiento de las primeras culturas LaTene a lo largo del río Marne (Francia), el rio Mosela (Alemania) y en Bohemia (República Checa).
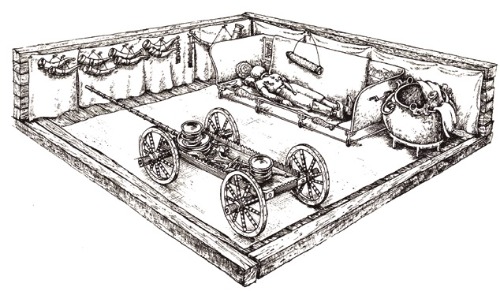
Una explosión demográfica alrededor del año 600 a.C. o tal vez luchas internas llevaron a miles de estas personas de Hallstatt D (y desde aproximadamente 400 a.C. a sus sucesores de LaTene) a cruzar los Alpes hasta llegar a las cálidas y acogedoras Tierras italianas. Otros fijarían sus ojos en territorios del este, instalándose en los Cárpatos en la antigua Yugoslavia, Hungría, Rumania y Bulgaria, y hasta Ucrania. Se dice que los celtas saquearon el santuario griego de Delfos en 279 a. C. y se establecieron en nuevos territorios en el sureste, particularmente en Anatolia. Algunos miembros del grupo anatolio volvieron sobre sus pasos para ocupar su antiguo hogar territorio y expandirse a nuevas regiones en el oeste cerca del Mar Mediterráneo, y al norte en las zonas belgas. Otros grupos se establecieron en zonas al oeste como España, al norte en Escandinavia.
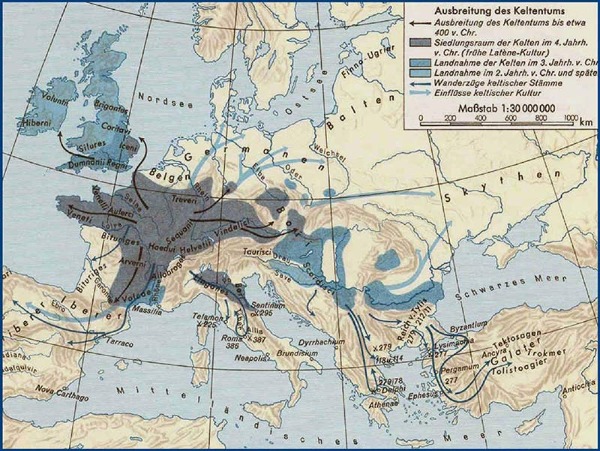
A ojos de los habitantes mediterráneos, los celtas fueron el prototipo pueblo “bárbaro” que, durante un tiempo, dominó la región entre la costa atlántica hasta las puertas de Asia en Anatolia y la moderna Ucrania. Desde la antigüedad “celta” se refiere en gran medida al pueblo conocido por este nombre (o variantes) por aquellos griegos y romanos, y que normalmente no incluían como tal a pueblos de habla celta de Gran Bretaña e Irlanda antes del año 100 a.C.
Estos autores o geógrafos greco-romanos empezaron a llamar “celtas” también a los habitantes de Masalia (Marsella), la zona mediterránea de la actual Francia anteriormente habitada por LIGURES e Iberos. Estrabón por ejemplo, en su libro IV, llama «celtas» a los instalados en los territorios opuestos a la Aquitania, a la zona del mar mediterráneo de Masalia y de Narbona, y que confinan con algunos montes de los Alpes (incluyendo Tolosa/Toulouse y su comarca claramente vasco-ibérica).
Genética celta centroeuropea. (Investigación del Y cromosoma R1b-U152, por David K. Faux)
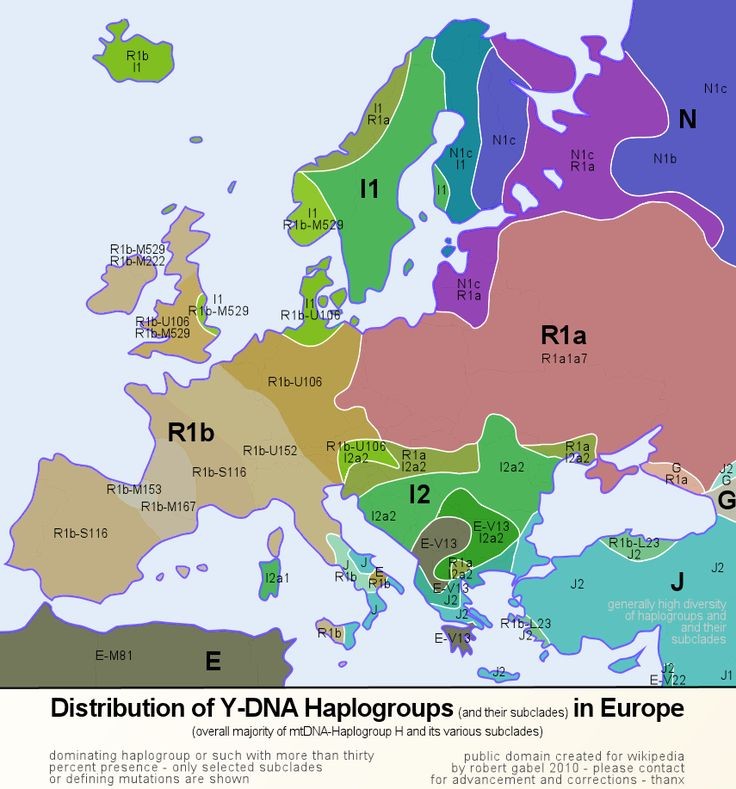
Últimamente se ha difundido la teoría del origen indoeuropeo de R1b-M269 en Europa, y se confunde a la población generalizando esta teoría a todo el grupo R1b. Olvidan que el hombre de Villabruna de Italia era de este grupo genético, un europeo R1b de hace 14.500 años. Se difunde esta errónea explicación, confundiendo al público, sin explicar la altísima frecuencia de dicho haplogrupo tanto entre los vascos o europeos occidentales actuales, como en las zonas más distantes de la presunta patria indoeuropea (Irlanda, y Highland), o en aquellas zonas donde vivieron (hasta la romanización) poblaciones pre-indoeuropeas como en Aquitania, el Levante español, Etruria, etc. En cambio, el haplogrupo R1b resulta casi ausente en la región que se presume es la patria de origen de las poblaciones indoeuropeas (Ucrania y las estepas euroasiáticas, cuya población es principalmente R1a).
Hasta la fecha, las teorías más aceptadas han argumentado que este patrón de frecuencias puede ser el resultado del origen y la posterior expansión posglaciar desde el refugio franco-cantábrico. La otra teoría, basada en la variación de los haplotipos Y-STR dentro de M269, también apoya su expansión posglacial, pero sostiene que M269 podría haber tenido una expansión paralela desde un refugio en Europa del Este (Anatolia). La nueva teoría propuesta por Balaresque, concluye que existe un origen único para el haplogrupo M269 en Europa del Este y desplaza el origen del período glacial al Neolítico, porque aplican tasas de mutación germinales más que evolutivas, generando tiempos de coalescencia más jóvenes.
La propuesta de Balaresque ha sido fuertemente refutada por Busby, que recalcularon la diversidad de haplotipos Y-STR dentro de M269 en una muestra más grande y geográficamente más amplia, lo que indicaba no una mayor diversidad en Europa del Este, sino un fondo homogéneo de variación de microsatélites en toda la muestra europea.
El DNA-Y tipo R-L51
R-L51 es típico de Europa occidental, es el verdadero haplotipo representativo del Atlántico y se habría originado en Europa central hace unos 6.000 años, aunque hay otra rama paralela entre los yamna. El R1b-L51es un clado intermedio entre los subgrupos L23 oriental y L11+ (P310+) occidental de R1b. Se encuentra en frecuencias extremadamente bajas desde Irán hasta Inglaterra, con una incidencia relativamente mayor en Europa central. Aún no está claro si surgió en Europa o en Anatolia, pero vale la pena mencionar que los subclados principales R1b P312 (U152, M167, L21) tienen su mayor variación en Francia, por lo tanto, se cree que L51 se formó en el sur de Francia (y no en Europa del este) durante el Neolítico tardío a partir de personasR1b L23 que habían llegado de Italia.
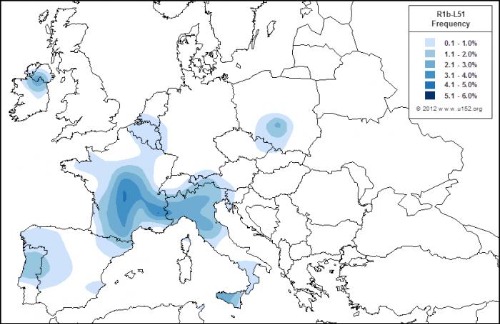
(mapa de R1b-L51 durante el neolítico. fuente Europedia)
Este mapa muestra altas frecuencias de este subclado en el sur/centro de Francia, así como en el norte de Italia. R1b L51 parece estar ausente de los Balcanes y Europa del este en general (excepto el sur de Polonia). Apuntar que L51 y su sucesor L11 se extendieron desde Europa occidental a Europa central y no al revés, lo que los hace no relacionados con las culturas de Europa central de la Edad del Bronce (Unetice,Urnfields, etc.).
Nuestro árbol filogenético del haplogrupo R1b muestra claramente que, además de la conocida rama R-L51 predominante en Europa occidental, existe otra rama R-GG400 distinta en Europa oriental. Los marcadores que definen esta rama oriental se revelaron en un conjunto de secuencias del cromosoma Y publicadas (Karminet al. 2015;Hal-last et al. 2015;Batini et al.2015). La rama oriental R-GG400 es de igual nivel filogenético y edad similar a la rama R-L51, pero con una distribución geográfica contrastante. La distribución de la rama occidental R-L51 es bien conocida, se encuentra en frecuencias más bajas en Europa del Este (Myres et al. 2011). Los datos recopilados sobre la distribución geográfica de la rama oriental de R-GG400 son escasos, pero ubican su área principal en las estepas de Europa del Este y Asia Occidental (Karmin et al. 2015) y muy escasa o nula presencia en el occidente europeo.
Las antiguas muestras Yamnaya se encuentran en la rama R-GG400 "oriental" del haplogrupo R1b-L23, lo que demuestra que los descendientes paternos de los Yamnaya todavía viven en la estepa póntica y que "LA ANTIGUA POBLACIÓN YAMNAYA NO FUE UNA FUENTE IMPORTANTE DE LINAJES PATERNOS EN LOSACTUALES EUROPEOS OCCIDENTALES", según Balanovsky, O.; Chukhryaeva, M.; Zaporozhchenko, V. del Instituto de Genética General Vavilov, Moscú, Rusia y del Centro de Investigación en Genética Médica,Moscú, Rusia. (2017, "LA DIFERENCIACIÓN GENÉTICA ENTRE POBLACIONES DE TIERRAS ALTAS YBAJAS DA FORMA AL PAISAJE CROMOSÓMICO Y DE ASIA OCCIDENTAL" (https://labs.icb.ufmg.br/lbem/pdf/Balanovsky2017HGlowlandAsia.pdf)
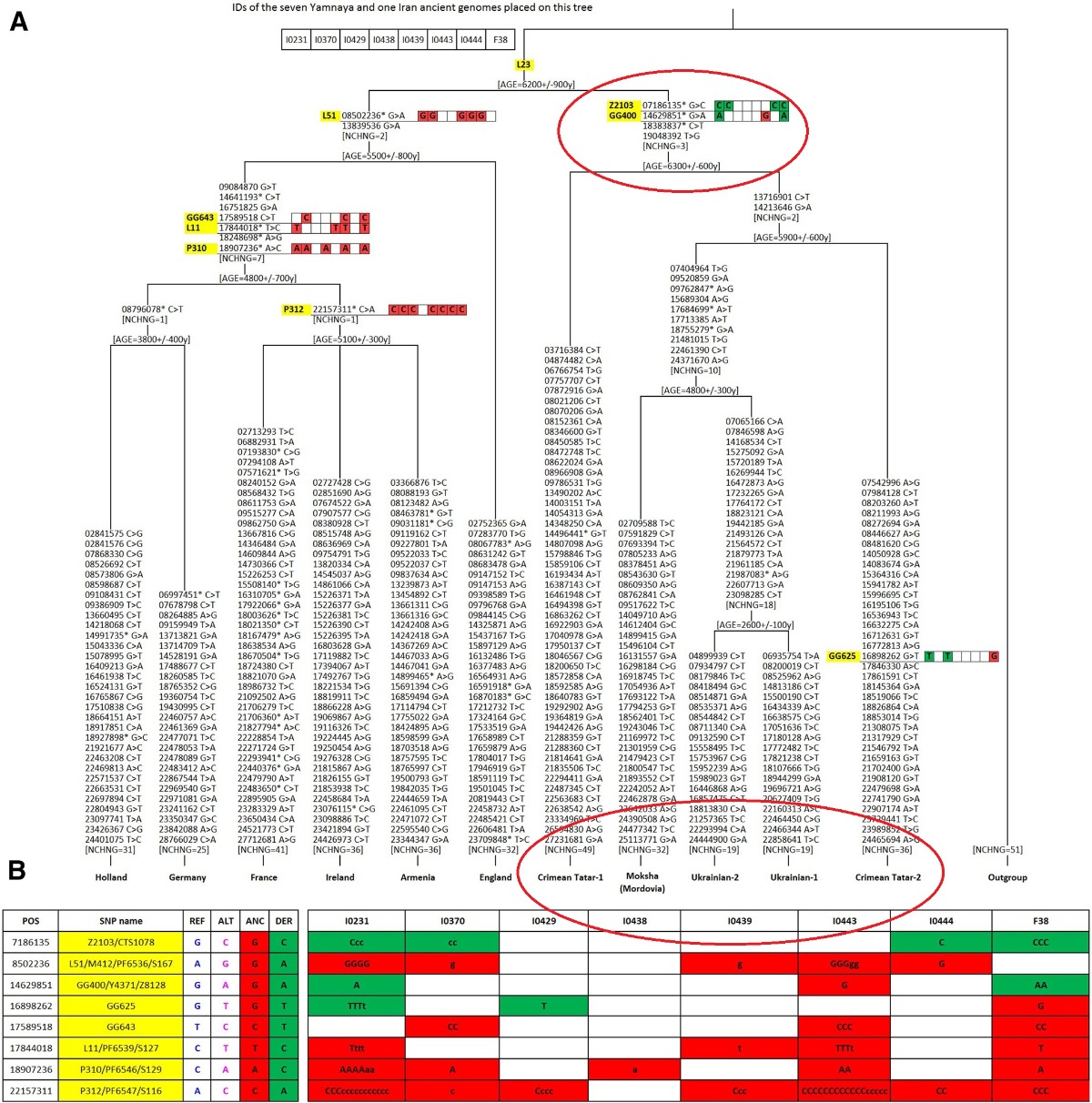
(PINCHA IMAGEN para agrandar)
David K. Faux estudio referencias históricas, datos lingüísticos y arqueológicos, para vincular los pueblos celtas Hallstatt y LaTene de Europa Central con un marcador de cromosoma Y, el U152 o S28 (se utiliza más el termino U152 debido a su aceptación por parte del Y Chromosome Consortium). La hipótesis que se está considerando es que la mayoría de los que son positivos para U152 son descendientes vivos de estos antiguos pueblos celtas que surgió en una población europea alpina. Con la excepción de gente lejanamente relacionada y que reside a lo largo de la costa mediterránea o al sur de Cerdeña.
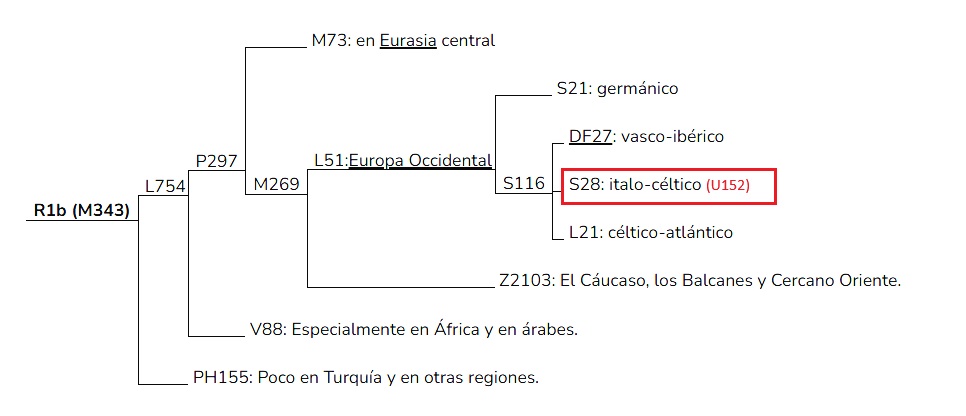
David K. Faux no propone que todos los celtas de Europa Central fueran U152, solo indica que hay una fuerte correlación (lo que no impide que también se dé una amplia gama de otros haplotipos encontrados entre los celtas). Para entender la dimensión de este parámetro genético se ha de poner como ejemplo su diversidad, como el cantante, compositor y bailarín estadounidense Michael Jackson (el Rey del Pop) que perteneció al haplogrupo R1b-U152, al igual que Abraham Lincoln o el actor Kevin Costner.
 ¿CUÁL ES EL ORIGEN DEL MARCADOR U152?
¿CUÁL ES EL ORIGEN DEL MARCADOR U152?
Todos los varones actuales que tienen el marcador U152 son descendientes de un hombre, que finalmente se convirtió en el progenitor de un gran porcentaje de las personas que serían conocidos como pertenecientes a LaTene celtica (sucesores de los anteriores pueblos de Hallstatt).
U152 es un solo polimorfismo de nucleótidos (SNP) desarrollado como S28 por Ethno Ancestry en 2005 (aún publicado de forma independiente en 2007 por Simms et al. quien llamó al marcador U152) como un subclado de M269 o R1b1b2* (y más recientemente se descubrió que está por debajo de P310, P311 (R1b1b2a*), y luego P312, o R1b1b1a2* de la cultura Megalítica atlántica y Bell Beaker (que sirve para unirse a un grupo haplogrupos como R-M167 y R-L21), y clado hermano de U106 (R1b1b2a1a).
Este U152 (también llamado polimorfismo de evento único o UEP) implica un punto Mutación en uno de los 60 millones de pares de bases de nucleótidos que componen el ADN del cromosoma Y. Pero se desconoce cuándo ocurrió este evento (las estimaciones oscilan entre 25.000 y Hace 2.500 años). Los antepasados de este hombre portaban el motivo RP312* (daría positivo para el estándar que define los marcadores M269 y P312, pero no el U152) en el cromosoma Y.
Sin embargo, cada uno de los descendientes directos de este individuo en línea masculina poseerá la misma mutación inofensiva pero informativa y ser asignada a la categoría filogenética de R1b1b2a2g* (Actualización YCC, 2008). Recientemente este haplogrupo se ha dividido en una forma ancestral (SNP L2 negativo) y una forma derivada más numerosa (L2 positivo), y un número menor derivado en L2 (S139) y positivo para el SNP L20 (S144).
La premisa del trabajo de David K. Faux es que, a pesar del tiempo trascurrido de 2500 años, se puede encontrar evidencia de la persistencia y supervivencia de Celtas centroeuropeos a través de un examen de la actual estructura del cromosoma Y en la población europea, con referencia específica al marcador U152. En resumen, David K. Faux postuló que el U152 se encontrará con frecuencia en áreas geográficas donde los datos históricos, lingüísticos y/o arqueológicos atestiguan la presencia de celtas La Tene (y Hallstatt), pero rara vez en otros lugares, a excepción de Cerdeña y la costa de Liguria y el sur de Italia (ya sea población aborigen o resultado de miles de esclavos galos importados a Italia en el siglo I a.C.).
En otras palabras, se debería contar que los varones que poseen la mutación U152 (o sus ancestros anteriores) representan un gran porcentaje de la población de Suiza. Por poco que sea, si es que hay alguno, debería observarse, por ejemplo, en el norte de los Países Bajos y el norte de Alemania, una región no documentada que tenga ningún vínculo con los celtas de Europa Central.
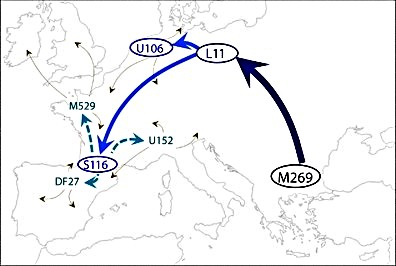
Aparentemente existen tres variedades principales de R-M269, según un YSNP recientemente descubierto, conocido como S116 o P312. Aquellos que tienen la versión ancestral incluyen aquellos con un haplotipo 35 “oriental” R-M269* tipo p49a,f (encontrado principalmente en Italia). Aquellos que dan positivo o derivan en P312 incluyen otros conocidos clados de R-M269, incluido el haplotipo 15 “occidental” R-M269*. Recientemente se ha descubierto que R-U106 es un clado hermano de R-P312.
A pesar de la superposición considerable, en general R-P312 se encuentra desde Europa Central hasta los puntos más lejanos confines más occidentales del continente, mientras que R-U106 se encuentra en Europa central hacia el norte hacia Escandinavia.
Generalmente se supone que todos estos grupos, con la excepción de R-M269* “oriental” Taq p49a,f el haplotipo 35, se retiraron al refugio glaciar Franco-Cantábrico en la LGM.
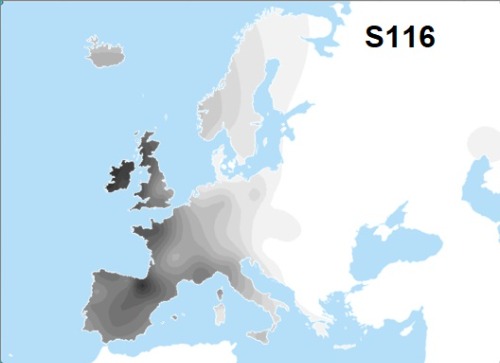
Considerando los patrones de distribución actuales, no sería sorprendente que gran parte del R-P312 permaneciera en la Península Ibérica durante y después de la LGM, mientras que el R-U106 puede haber sido asociado con el Refugio de los Balcanes.
Sin embargo, en última instancia, algunos R-P312* se abrieron camino, tal vez a lo largo del margen costero del norte de España o al oeste del Macizo Central de los franceses, por la costa mediterránea hasta el norte de Francia. Quizás R-U152 (o sus ancestros R-P312*), refiriéndose nuevamente a la distribución que se observa hoy, siguió un camino al este del Macizo Central, a lo largo del corredor Ródano-Saona, luego hacia el este siguiendo el margen glaciar alpino hasta las cabeceras de los ríos Rin y Danubio en Suiza y el sur de Alemania.
La región de las cabeceras del Rin y el Danubio experimenta un aumento significativo en el número de poblados entre 16mil y 14,7mil años antes del presente. Durante el Magdaleniense posterior se encuentran asentamientos donde se ven grandes campamentos al aire libre en el lago Neuchatel, en las cuencas de Neuweid y París, y estos se corresponden con importantes ocupaciones de refugios rocosos en la cuenca del Rin-Danubio, las tierras altas de Sur de Alemania, Turingia y Bélgica. Estas áreas están destinadas a convertirse en futuros territorios de origen de los celtas. Este patrón sería consistente con lo que se sabe de la cultura magdaleniense, con un patrón de distribución de utensilios que refleja lo que podría ser propuesto en relación con la expansión de U152 desde el refugio de la Aquitania.
Los mapas con sitios arqueológicos que datan del Magdaleniense superior (Bocquet-Appel, 2005) muestran fuerte agrupación en las áreas habitadas durante la época LGM (por ejemplo, el sur de Francia), pero ahora también en una franja que se extiende de suroeste a noreste con poca actividad en zonas del sur, excepto alrededor del Mar Negro. Un número significativo de sitios se ven a lo largo de la Corredor del Ródano con los dos mayores grupos/agrupaciones periféricas en la zona que hoy son los Países Bajos y el norte de Alemania, que corresponden a Hamburgo-Grupo Aherensberg (R-U106); y el Distrito Suizo de los Lagos más suroeste de Alemania (R-U152).
El Magdaleniense, abarcó una vasta área en su apogeo, y es totalmente consistente con la distribución conocida de R-M269 y los clados actuales (con una caída importante en el número de personas desde la actual Polonia hacia el este).
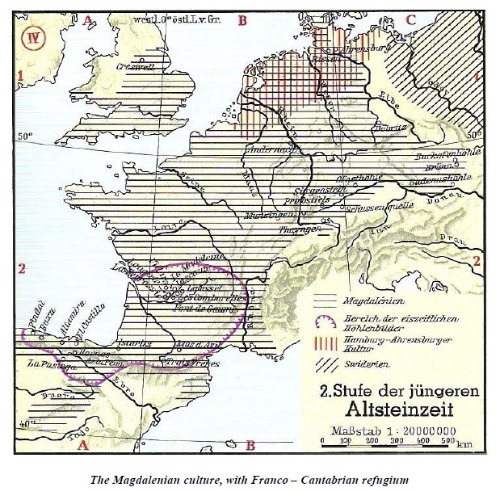
LA CULTURA MAGDALENIENSE, CON EL REFUGIO FRANCO-CANTÁBRICO
El mapa superior muestra la ubicación del refugio Franco-Cantábrico dentro de la zona. rodeado por la línea violeta, e ilustra cómo el Magdaleniense (representado por líneas horizontales) incluyeron en última instancia prácticamente toda Europa occidental al norte de los Alpes, pero Terminaba entre los ríos Elba y Danubio.
La migración de humanos desde la zona oriental del refugio franco-cantábrico Aquitania podría haber incluido a los antepasados del R-U152 que seguirían el corredor del Ródano al noreste, como se muestra en el mapa a continuación. Hay dos clados de R-U152, uno con el corto estable marcador de repetición en tándem (STR) DYS492=12 (modal para R-M269*) y un marcador más pequeño porcentaje con 14 repeticiones. Si bien sólo las pruebas de ADN antiguo proporcionarán una respuesta inequívoca, una hipótesis es que esta primera ola de U152, durante época magdaleniense, era principal o exclusivamente del menos común DYS492=14 (que es la forma ancestral, L2 negativa) variedad, y que aquellos cuyo haplotipo incluye DYS492=14 seguirían el mismo camino hacia el norte que sus primos lejanos, o quizás a través de una doble migración desde la costa mediterránea hacia el norte a través de los Alpes y una ruta más tortuosa en semicírculo hacia el oeste y luego hacia el norte para evitar los Alpes.
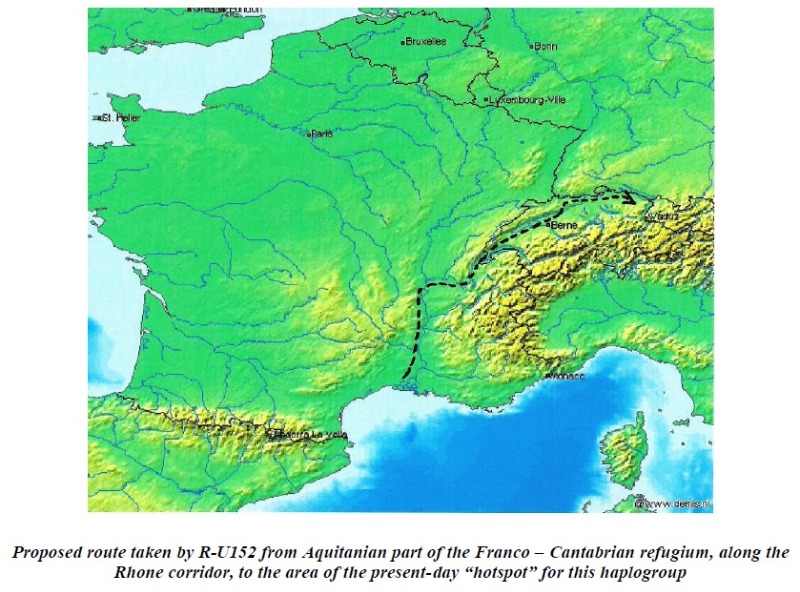
Esta propuesta es probable que el movimiento no haya ocurrido hasta los primeros años del Neolítico.
Bibliografía:
• New clues to the evolutionary history of the main European paternal lineage M269: dissection of the Y-SNP S116 in Atlantic Europe and Iberia. https://www.nature.com/articles/ejhg2015114#citeas
• AGeneticSignalofCentralEuropeanCelticAncestry: PreliminaryResearchConcerningY-Chromosome MarkerU152. DavidK.Faux. https://www.davidkfaux.org/LaTene_Celt_R1b1c10.pdf
• https://www.kondaira.net/esp/Euskara0003.html