
 Las evidencias actuales demuestran que la persistencia a la lactasa, la encima que nos permite beber leche, NO es por la llegada de hablantes indoeuropeos de la estepa póntica en la Edad del Bronce. Los pastores de Asia y los individuos de los kurganes no disponen de alelo T*13910.
Las evidencias actuales demuestran que la persistencia a la lactasa, la encima que nos permite beber leche, NO es por la llegada de hablantes indoeuropeos de la estepa póntica en la Edad del Bronce. Los pastores de Asia y los individuos de los kurganes no disponen de alelo T*13910.
La lactasa, que se encuentra principalmente en las células del intestino delgado de los recién nacidos y algunos adultos, es la enzima encargada de descomponer la lactosa. Su ausencia provoca síntomas de intolerancia a la lactosa en quienes consumen productos lácteos.
Después del período de lactancia, los niveles de lactasa disminuyen en la mayoría de los mamíferos, incluido el ser humano. Esta reducción enzimática impide la correcta digestión de la lactosa, lo que provoca molestias en los adultos que consumen leche.
A pesar de la disminución generalizada de la lactasa, algunas personas mantienen la enzima activa durante toda su vida. Este fenómeno se debe a mutaciones genéticas que permiten la persistencia de la lactasa, permitiendo así el consumo de leche sin efectos negativos. El desarrollo de la persistencia de la lactasa está vinculado a la historia evolutiva, particularmente a la revolución agrícola, el pastoreo y el consumo de productos lácteos. En regiones como el norte de Europa, la lactasa persistente ofreció una ventaja evolutiva, ya que la lactosa ayudaba en la absorción del calcio, crucial para la salud ósea en zonas de baja radiación solar.
El consumo de leche ofrece una ventaja nutricional, su contenido en ácido p-aminobenzoico es inferior al de las dietas no lácteas y en su composición se encuentran inmunomoduladores. Relacionado está por lo tanto el estudio que sugirió en el que la persistencia de la lactasa se seleccionó de forma paralela a la resistencia a la malaria en los fulani (Malí).
Variantes Genéticas Asociadas a la Persistencia de la Lactasa
Se han identificado varios marcadores genéticos de la persistencia de la lactasa que muestran que esta tiene múltiples orígenes en diferentes partes del mundo (ejemplo de evolución convergente).
Dentro de Europa y en las poblaciones de ascendencia europea, están casi totalmente correlacionados con la presencia de la mutación C/T*13910 (rs4988235). Aunque también se encuentra en Asia central, Asia sur, y pequeñas poblaciones de Mali, Sudán y Camerún.
La frecuencia de esta persistencia varía mucho según la región: es alta en las poblaciones del norte de Europa (>90% en Suecia y Dinamarca), en la población moderna del País Vasco es el 66%, mientras que disminuye progresivamente hacia el sur de Europa y Oriente Medio (alrededor del 50% en Francia, España y algunas poblaciones árabes), y es muy baja en las poblaciones asiáticas y africanas, aunque es común en las poblaciones pastoriles (alrededor del 1% en China, 5-20% en África Oriental).
Sin embargo, este polimorfismo no es un buen indicador de la persistencia de lactasa en los no europeos, ya que las poblaciones de pastores de África y Asia, aunque muestran una alta prevalencia de persistencia de lactasa, presentan una baja frecuencia del alelo T*13910.
Esta homogeneidad europea difiere de las distribuciones alélicas de la persistencia de la lactasa en el resto del mundo, especialmente en África y Oriente Medio, donde coexisten varios alelos:
- T/G*13915 está presente sobre todo en poblaciones del este y norte de África y Oriente Medio.
- G/C*14010 se identificó en África oriental.
- C/G*13907 se describió en Sudán y Etiopía
Para estimar la edad de los SNP asociados con la capacidad de digerir la lactosa los investigadores han utilizado diferentes métodos computacionales. En el caso de Europa, la edad del alelo T*13910, se ha estimado mediante dos métodos basados en la acumulación de microsatélites; el programa Ytime (Behar et al., 2003) y el método que muestra la reducción en la frecuencia de microsatélites asociados con el alelo T*13910 (Seixas et al., 2001):
- El programa Ytime (Behar et al., 2003): el alelo surgió hace entre 4125 y 11000 años.
- El método de recombinación data 7500 a 3000 años (Coelho et al., 2005).
Porcentaje de adultos con un genotipo T*13910 (rs4988235) de persistencia de la lactasa en la población antigua del Viejo Mundo.
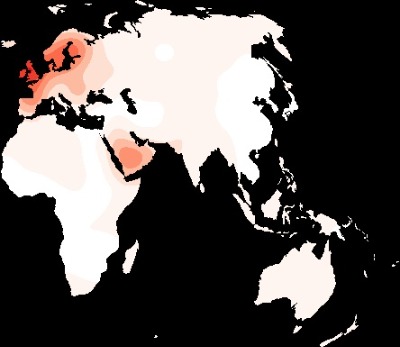
Mapa del porcentaje en la población adulta indígena del Viejo Mundo. Genotipos conocidos de persistencia de la lactasa (-14010 G>C, -13915 T>G, -13907 C>G o -13910 C>T). Los círculos marcan las ubicaciones de las muestras.
Se ha planteado la hipótesis de que la variante T*13910 apareció al menos dos veces de forma independiente, observándose en dos haplotipos diferentes. El H98 es el más común, hallándose en los finlandeses y los fulani, los H8 y H12 están relacionados con poblaciones geográficamente restringidas.
Los intentos de establecer cuándo y dónde comenzó la selección del alelo T*13910 se basan hasta ahora en una hipótesis anticuada que propone a la llanura húngara/cuenca de los Cárpatos y aproximadamente hace unos 7.500 años. Creen erróneamente que T*13910 fue llevado por los primeros agricultores, muy probablemente por la gente LBK, aunque ninguno de los individuos de esta cultura, estudiados hasta ahora, tenían este alelo T*13910.
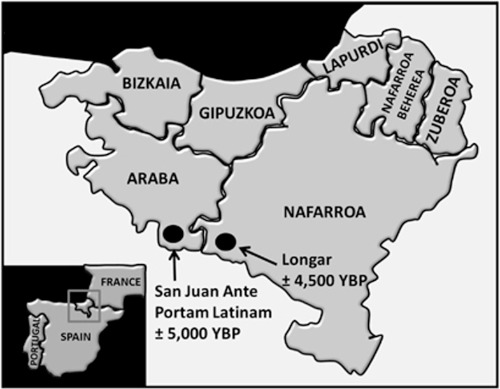
Las únicas personas neolíticas persistentes en la lactasa encontradas hasta ahora vivieron mucho después de que la gente LBK entrara en Europa Central, es decir, entre 5.000 y 4.500 años, en poblaciones que representan diversas culturas arqueológicas. Se identificaron entre los individuos del Neolítico Tardío/Calcolítico, dos individuos que vivían en el territorio del País Vasco (T = 0,23; LP = 0,27) y en Escandinavia aparece el alelo sorprendentemente entre cazadores-recolectores tardíos de la cultura de la Cerámica Picada (CPC) (T y LP =0,05) [ 67 ].
¿Por qué no se encontró el alelo T en los agricultores que entraron en Europa 7.500 años antes del presente?
Estos agricultores tenían acceso a la leche, el principal factor necesario en el proceso de selección, pero aparentemente tener acceso a la leche no era lo más importante. Es sorprendente que el alelo T*13910 no se haya encontrado, hasta ahora, ni a lo largo de la ruta del Danubio ni del Mediterráneo. Esta carencia puede ser debido a que:
- Estaba presente con una frecuencia muy baja, lo que podría indicar que las personas LBK no consumían leche fresca.
- No estaba presente en absoluto entre los primeros agricultores (culturas LBK y Cardinal), no siendo de origen neolítico temprano, ya que se encontró en Iberia solo aproximadamente 5.000 años, y 4.500 años en personas cazadores-recolectores que vivían en la isla Gotland del mar báltico [ 67 ].
La falta observada del alelo T*13910 en las primeras poblaciones neolíticas, así como en las posteriores, indica su origen bastante distinto al de las personas LBK. El alelo T*13910 no fue encontrado entre las personas LBK ni en las posteriores a LBK, es decir, pertenecientes a las culturas Lengyel y Ánfora globular, y aunque los primeros agricultores tuvieron la oportunidad de transmitirlo durante al menos 6 siglos este alelo no fue trasmitido.
Dos rutas son las principales en el proceso de neolitización: una a lo largo del río Danubio, que conduce a Europa central, y la otra a lo largo de la costa norte del mar Mediterráneo.
El alelo T*13910 NO SE ENCONTRÓ en los primeros agricultores europeos de esas dos rutas tradicionales de neolitización, seis de los cuales fueron enterrados en la llanura húngara y en Alemania central. Se encontraron esqueletos de personas relacionados con la Cultura Cardial de Trielle al sur de Francia, un sitio ubicado en la ruta neolítica mediterránea de hace 5.000 años, que dieron negativo al T*13910. Casi de la misma edad, aproximadamente 600 km al oeste y en el área del País Vasco moderno, vivía una población que hasta el 27% era tolerante a la lactosa [66].
¿Pudo haberse expandido el alelo T*13910 a Europa por otra ruta?
Si el alelo no se transmitió por las dos vías neolíticas principales, podría haber estado ya presente en la población mesolítica ancestral de los vascos modernos. Podría haber sido introducido en Iberia, por ejemplo, por personas procedentes de África a través del Estrecho de Gibraltar.
La alta frecuencia observada en el área del País Vasco hace 5.000–4.500 años [66], en contraste con Europa central [14] y meridional [13], justifica una hipótesis sobre un escenario de propagación distinto del alelo T*13910 (Fig. 5). Este podría haberse propagado a Europa a lo largo de una Ruta Norte (NR), que corre hacia el este desde el norte de Iberia a lo largo de la costa europea, por mar y/o por tierra, llegando principalmente a las regiones del norte donde hoy se observa una alta frecuencia de T*13910.
La Ruta Norte marina podría haber corrido a lo largo del Golfo de Vizcaya, la bretaña francesa, el Canal de la Mancha hasta el Mar del Norte y a través de Skagerrak y Kattegat hasta el Mar Báltico, "entregando" el alelo no solo a la parte norte de Europa continental, sino también a los archipiélagos del noroeste de Europa.
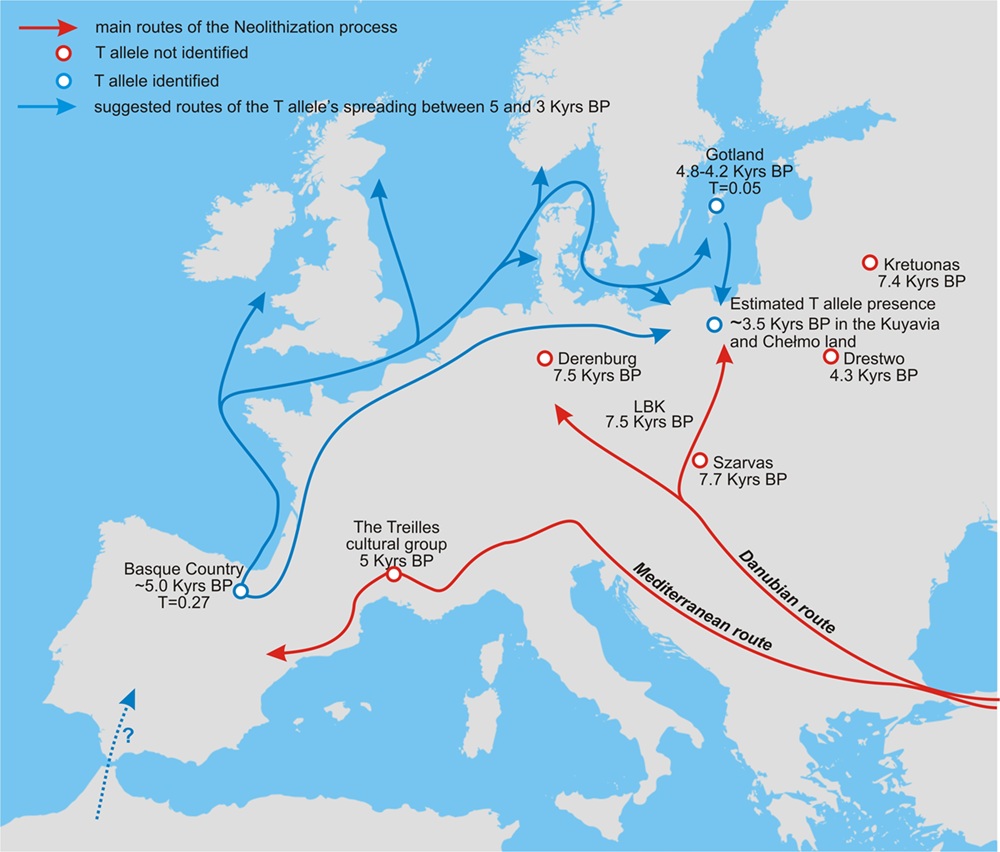
(Fig. 5)
Se descubrió que los primeros agricultores llegaron al norte de Gran Bretaña, las Islas del Canal y la Isla de Man hace 5.700 años. Tomando el escenario sugerido de la ruta de propagación del alelo T a lo largo de la Ruta del Norte desde Iberia, donde se atestigua su presencia hace 5.000 años en el País Vasco, es posible que el alelo llegara a las británicas con ganaderos-agricultores ibéricos. Según Olalde, los individuos neolíticos del sur de Francia y Gran Bretaña están significativamente más cerca de los agricultores ibéricos del Neolítico Temprano que de los agricultores centroeuropeos del mismo periodo. [4]
Siguiendo esta ruta hacia el mar Báltico, la población cazadora-recolectora de Gotland (PWC), habitó la isla simultáneamente con otra población neolítica NO INDOEUROPEA que practicaba la producción lechera desde hace 6.000-5.000 años [69] relacionada con el llamado Vaso de Embudo (TRB). El hallazgo del alelo T*13910 en las personas de Gotland que vivieron entre 4.800 y 4.200 años, así como en las de Polonia central (que vivieron aproximadamente entre 4.000 y 3.000 años), podrían estar expuestas al alelo T de una forma diferente de los individuos de Europa central y meridional. La ruta marítima del norte en la propagación del alelo es aún más probable a la luz de la similitud genética sugerida recientemente entre los mesolíticos (Lola) y agricultores escandinavos con respecto de los ibéricos [70].
Lo que está claro, con las evidencias actuales, es que la persistencia a la lactasa en de las culturas de la Edad del Bronce no es por la llegada de hablantes indoeuropeos de la estepa póntica. Los pastores de Asia, presentan una baja frecuencia del alelo T*13910 y los individuos de los kurganes no disponen de alelo T*13910.
Un estudio genético publicado en noviembre de 2020 por Joachim Burger et al. analizaron el alelo T*13910 en muestras genéticas antiguas, tomaron 14 muestras de la masacre de Tollense (Alemania, 1250 a. C.) y las cruzaron con otras anteriores a la edad del Bronce de Serbia, Europa del Este y Estepa póntico-caspia llegando a la siguiente conclusión: <<Inferimos una baja LP (persistencia lactasa) en las tres regiones, es decir, en el norte de Alemania y el sureste y este de Europa, lo que sugiere que el aumento de rs4988235 (alelo T*13910) en Europa central y septentrional probablemente no fue causado por expansiones esteparias.>> [10]
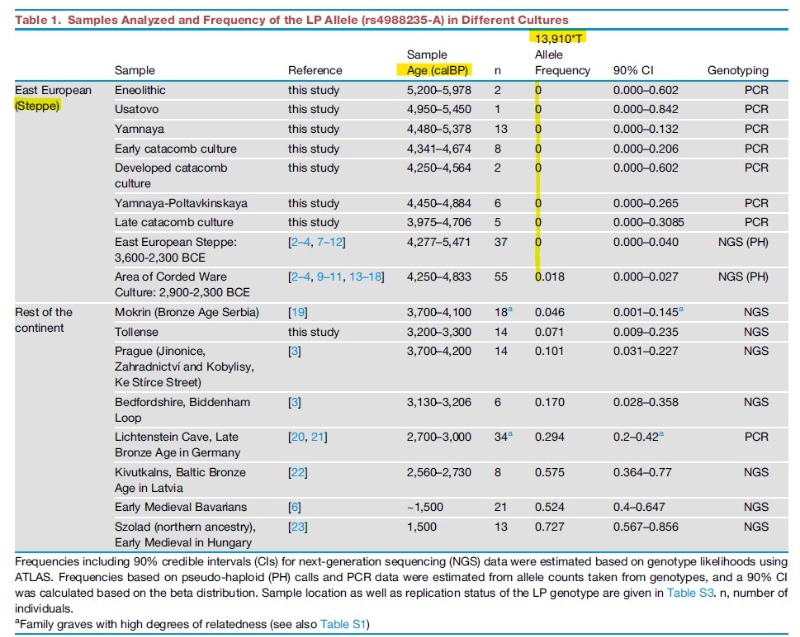
Este estudio de paleo genética de Burger realizado con MUESTRAS REALES es una crítica de los análisis genéticos computacionales, como los realizados en 2009 por Yuval Itán, Adán Powell et al. publicados en LOS ORÍGENES DE LA PERSISTENCIA DE LA LACTASA EN EUROPA, y que se basaban en un cálculo matemático entre frecuencia actual del alelo T*13910, las fechas de llegada de la agricultura a Europa mediante un cálculo bayesiano aproximado.
Casualmente se encontró en la cueva de Lichtenstein en Alemania una frecuencia del 27% (ver Schilz 2006 ) del alelo T*13910 entre 13 individuos del Bronce Final la cultura Campos Urnas (1.250-750 a.C.), porcentaje similar a la de Iberia, que también localiza un 27% de frecuencia en el área del País Vasco hace 5.000 años, y 2.000 años antes de que llegaran los Campos Urnas a la zona del alto Ebro.
Se cree que los agricultores neolíticos del noreste de África y Oriente Próximo, pueden ser la población de origen de la variante T* 13910 y que posteriormente hayan sido suplantadas por migraciones posteriores de pueblos. Los fulani del África occidental subsahariana, los tuaregs del norte de África y los agricultores europeos descendientes de estos agricultores neolíticos pueden por ello compartir esta variante de persistencia de la lactasa. Tishkoff y col. (2017) confirmaron que los hausa y los fulani, dos tribus del Sahel con alta incidencia de R1b-V88, poseían el mismo alelo T* 13910 que los europeos.
Mediante el análisis de grasas halladas en fragmentos de cerámica, se ha demostrado que la producción de leche comenzó en la llanura panónica de Hungría hace 7.000 años (Craig et al., 2005) y, aproximadamente cien años después, en Gran Bretaña (Copley et al., 2003).
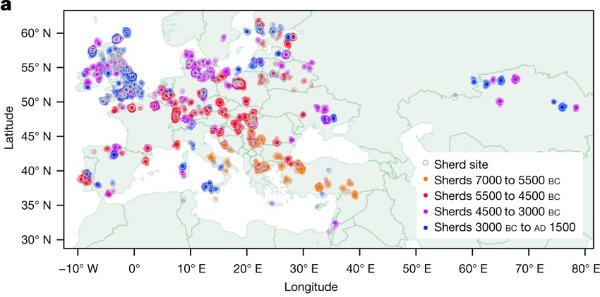 Imagen a : Los círculos de colores ilustran 6.899 cerámicas con grasas animales de 826 fases de 554 yacimientos (círculos grises). Los círculos de colores están ligeramente movidos para evitar una superposición, lo que permite visualizar el número de tiestos.
Imagen a : Los círculos de colores ilustran 6.899 cerámicas con grasas animales de 826 fases de 554 yacimientos (círculos grises). Los círculos de colores están ligeramente movidos para evitar una superposición, lo que permite visualizar el número de tiestos.
En cuanto a África, sabemos que el Neolítico se desarrolló más tarde. Otros análisis de grasas halladas en fragmentos de cerámica en África han indicado que las poblaciones saharianas comenzaron a consumir leche hace 5.000 años (Dunne et al., 2012).
Panorámica del yacimiento de El Portalón de Cueva Mayor, en la sierra de Atapuerca, del que se han recuperado cerámicas con restos de grasas lácteas y en el estudio de los restos humanos de Portalón en 2014, los individuos no eran tolerantes a la lactosa. El individuo del Portalon de 5.373 años y código de identificación ATP3 tenía una Y-DNA tipo R1b-M173>V88 (LipsonNature2017). A escasos kilómetros si tenemos la persistencia de lactasa o alelo T* 13910 que apareció hace 5.000 años por primera vez en Alaba, aunque se mantuvo en baja frecuencia hasta hace 3.000 años, en la Edad del Bronce, en la que ya presenta frecuencias muy apreciables.
En apoyo de esto, un estudio en la Península Ibérica también ha observado que el alelo -13910 T aparece con menor frecuencia en poblaciones antiguas que en poblaciones modernas (Plantinga et al., 2012). Parece que, cuando el Neolítico llegó a Europa, el número de individuos resistentes a la lactasa era muy bajo. Por lo tanto, si la mutación estaba presente en la población, y debido a los beneficios que ofrecía el consumo de leche, la selección natural habría favorecido al alelo resistente a la lactasa.
Se han analizado muchas muestras antiguas de ADN del Mesolítico, Neolítico y la Edad del Bronce en Europa y Siria, y el alelo T* 13910 solo se ha encontrado en individuos del Neolítico Tardío/Calcolítico y de la Edad del Bronce. Los estudios de ADNa en Europa Central han demostrado que las poblaciones agrícolas del Neolítico temprano aún no se habían adaptado al consumo de leche, de hecho, no se ha detectado el alelo T* 13910 en muestras de ADN antiguo (Burger et al., 2007).
Para explicar las diferencias entre las poblaciones antiguas y modernas, se ha propuesto que los ancestros de algunas poblaciones europeas modernas fueron reemplazados por poblaciones agrícolas y ganaderas (Malmström et al., 2010). Un reemplazo por «selección natural turboalimentada» como explica Mark Thomas (University College of London) entre autóctonos y no por inmigración.
Es lo que tiene más sentido, por ejemplo en las islas Británicas, donde según cita Olalde[4], los análisis de isótopos de estroncio y oxígeno de esqueletos británicos de los períodos campaniforme y de la Edad del Bronce, NO HAN APORTADO EVIDENCIA DE MOVILIDAD sustancial a lo largo de la vida de los individuos desde lugares con climas más fríos o con geologías atípicas de Gran Bretaña. Por ello el reemplazo británico es por “selección natural” entre los propios británicos. Que conocen la agricultura y la cultura del vaso campaniforme en el 2500 a. C. en un pico cálido, pero posteriormente sufren una brutal bajada de la temperatura mundial que duró 1.000 años, del 2.300 hasta el 1.300 a.C. arruinando la agricultura y dejando la supervivencia en manos de los ganaderos que ya ordeñaban por aquel entonces en Gran Bretaña.
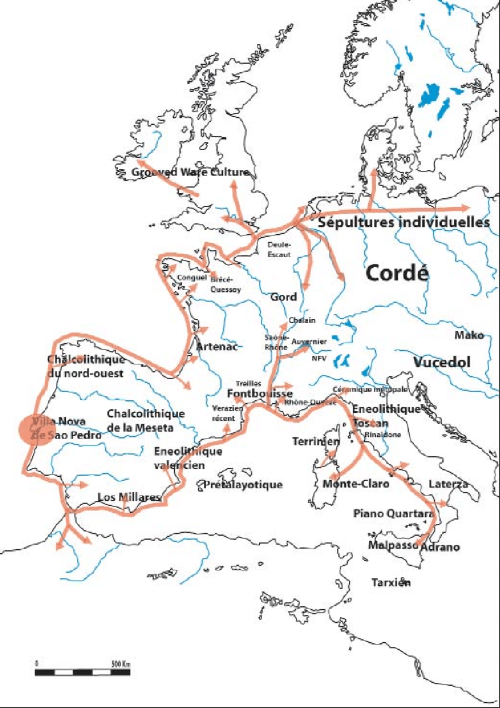
(imagen superior, rutas del campaniforme marítimo)
Un estudio arqueo-genético de 2021, descubrió que la persistencia de la lactasa aumentó rápidamente en Gran Bretaña en los inicios de la Edad del Hierro, mil años antes de que se generalizara en la Europa continental, lo que sugiere que la leche se convirtió en un alimento básico o muy importante en Gran Bretaña en esa época. Actualmente, la persistencia a la lactasa es aproximadamente proporcional al porcentaje de R1b, y en menor medida al de R1a, presente en una población. En las Islas Británicas, los Países Bajos y el suroeste de Escandinavia, donde el LP (persistencia a la lactasa) es el más alto del mundo, el porcentaje combinado de R1a y R1b supera el 70% de la población. En Iberia, el mayor porcentaje de LP se observa entre los vascos, que presentan también el mayor porcentaje de R1b. En Italia, el LP es más común en el norte, al igual que el R1b. La menor incidencia de LP en Europa se encuentra en el sur de Italia, Grecia y los Balcanes, las regiones con menos linajes de R1b.
¿Cuál es el origen del alelo de persistencia de la lactasa en los europeos?
Los investigadores concluyen que son las economías de subsistencia y la exposición a patógenos los fatores que mejor pueden explicar la persistencia de la lactasa en Europa.
Se puede especular que, en poblaciones productoras de leche y ganaderas, tanto la alta mortalidad como la fertilidad podrían haber influido en la tasa de selección de alelos LP, lo que resultó en una mayor tasa de supervivencia de los niños post-destete con persistencia de la lactasa, un efecto pronunciado en condiciones de vida difíciles. Dicho mecanismo podría modular el proceso de selección de alelos de persistencia en la lactasa tanto en la fría Europa del Norte como en las regiones cálidas y áridas del noroeste de África y la Península Arábiga. De hecho, los agentes que influyen en la mortalidad de los niños post-destete, a través del acceso restringido a alimentos y agua en la población que practica la cría de ganado y el procesamiento de leche, deberían considerarse presumiblemente involucrados en la selección de estos alelos de persistencia a la lactasa.
Planteamos la hipótesis de que ningún agente en particular estuvo involucrado en el proceso de selección del alelo T además de las necesidades básicas de vida relacionadas con la supervivencia.
Aunque beber leche por adultos intolerantes a la lactosa no es potencialmente mortal, esta incapacidad en los niños después del destete, que enfrentan hambre y sed, podría haber sido motivo de alta mortalidad en dichos niños, podría haber resultado principalmente de la desnutrición y la diarrea junto con las infecciones que son comunes incluso hoy en día, probablemente favoreció la supervivencia de los niños tolerantes a la lactosa que tenían acceso a la leche disponible en aquellas poblaciones neolíticas y post-neolíticas.
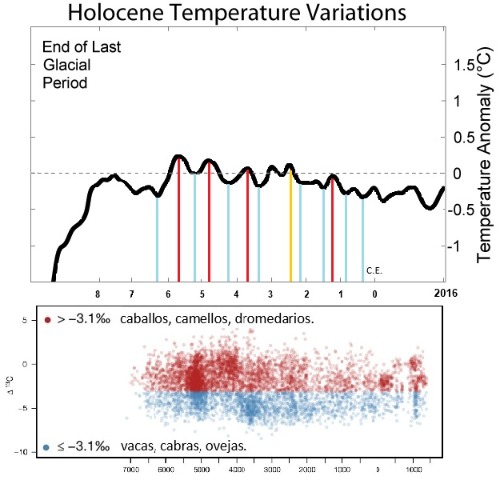
Gráfico superior de valores de Δ¹³C para 6.899 residuos de grasa animal extraídos de cerámicas prehistóricos a lo largo del tiempo. Cada punto es una medición individual, con su posición en el eje x muestreada aleatoriamente del rango de fechas uniforme de la fase arqueológica asociada. Las grasas animales con valores de Δ¹³C ≤−3,1‰ se definen como grasas de leche de rumiantes (vacas, cabras, ovejas) en color AZUL. La leche de yegua no se caracteriza utilizando este proxy⁸⁴ y, por lo tanto, se definiría como >−3,1‰. en color ROJO.
Además de la alta mortalidad, la alta fertilidad podría convertirse en la segunda de las fuerzas impulsoras que alimentaron el proceso de selección del alelo T de forma natural.
Por lo tanto, la introducción del alelo en cualquier población lechera tuvo que resultar en el comienzo de sus cambios de frecuencia. Sin duda, las condiciones ambientales desventajosas y cambiantes que afectaron a los cultivos, como se sugirió para el Neolítico, que probablemente resultaron en escasez de alimentos, podrían haber estado involucradas en la estimulación de la tasa de selección. Por lo tanto, cuanto más al norte se encontraba un área habitada y cuanto más duras eran las condiciones de vida, más fuerte era la presión de selección que operaba a través de tasas de mortalidad/fertilidad alterada.
La tolerancia a la lactosa, es un fenómeno que los expertos consideran una «selección natural turboalimentada», explica Mark Thomas, investigador del University College of London (UCL).
Redactado por I. Leibar
Bibliografia:
04- Iñigo Olalde, et al.The Beaker phenomenon and the genomic transformation of northwest Europe. N A T U R E | 2 0 1 8. doi:10.1038/nature25738
05- Theo S Plantinga, Santos Alonso, Neskuts Izagirre, Montserrat Hervella, Rosa Fregel, Jos WM van der Meer, Mihai G Netea, Concepcion de la Rúa. Low prevalence of lactase persistence in Neolithic South-West Europe. https://pmc.ncbi.nlm.nih.gov/articles/PMC3376259/
07- Burger J, Kirchner M, Bramanti B, Haak W, Thomas MG. Absence of the lactase-persistence-associated allele in early Neolithic Europeans. Proc Natl Acad Sci U S A. 2007 Mar 6;104(10):3736-41. doi: 10.1073/pnas.0607187104. Epub 2007 Feb 28. PMID: 17360422; PMCID: PMC1820653. https://pubmed.ncbi.nlm.nih.gov/17360422/
08- Henryk W. Witas , Tomasz Płoszaj, Krystyna Jędrychowska-Dańska, Piotr J. Witas, Alicja Masłowska, Blandyna Jerszyńska, Tomasz Kozłowski, Grzegorz Osipowicz. Hunting for the LCT-13910*T Allele between the Middle Neolithic and the Middle Ages Suggests Its Absence in Dairying LBK People Entering the Kuyavia Region in the 8th Millennium BP. Published: April 8, 2015 https://doi.org/10.1371/journal.pone.0122384.
09- Jose A. Peña, Miguel A. Alfonso-Sánchez, Susana García-Obregón, Ana Pérez-Miranda. Lactase persistence in residing population of the Basque Country. http://www.didac.ehu.es/antropo/3/3-4/Pena.pdf
10- Joachim Burger, Vivian Link, Jens Blöcher, Anna Schulz, Christian Sell, Zoé Pochon, Yoan Diekmann, Aleksandra Žegarac, Zuzana Hofmanová, Laura Winkelbach, Carlos S. Reyna-Blanco, Vanessa Bieker, Jörg Orschiedt, Ute Brinker, Amelie Scheu, Christoph Leuenberger, Thomas S. Bertino, Ruth Bollongino, Gundula Lidke, Sofija Stefanović, Detlef Jantzen, Elke Kaiser, Thomas Terberger, Mark G. Thomas, Krishna R. Veeramah, Daniel Wegmann. Low Prevalence of Lactase Persistence in Bronze Age Europe Indicates Ongoing Strong Selection over the Last 3,000 Years, Current Biology, Volume 30, Issue 21, 2020, Pages 4307-4315.e13, ISSN 0960-9822, https://doi.org/10.1016/j.cub.2020.08.033.
11- Yuval Itán,Adán Powell,Mark A. Beaumont,Joachim Burger,Mark G. Thomas. Los orígenes de la persistencia de la lactasa en Europa. Publicado: 28 de agosto de 2009. https://doi.org/10.1371/journal.pcbi.1000491
13- Lacan M, Keyser C, Ricaut FX, Brucato N, Duranthon F, Guilaine J, et al. (2011) El ADN antiguo revela la difusión masculina a través de la rutamediterránea neolítica. Proc Nat Acad Sci USA 108: 9788–9791. pmid:21628562 https://doi.org/10.1073/pnas.1100723108
14- Burger J, Kirchner M, Bramanti B, Haak W, Thomas MG (2007) Ausencia del alelo asociado a la persistencia de la lactasa en los primeros europeos del Neolítico. Proc Natl Acad Sci USA 104: 3736–3741. pmid:17360422 https://doi.org/10.1073/pnas.0607187104
66-Plantinga TS, Alonso S, Izagirre N, Hervella M, Fregel R, van der Meer JW, et al. (2012) Baja prevalencia de persistencia de lactasa en el suroeste de Europa neolítica. Eur J Hum Genet 20: 778–782. pmd:22234158 https://pubmed.ncbi.nlm.nih.gov/22234158/
67- Malmström, H., Linderholm, A., Lidén, K. et al. High frequency of lactose intolerance in a prehistoric hunter-gatherer population in northern Europe. BMC Evol Biol 10, 89 (2010). https://doi.org/10.1186/1471-2148-10-89
70- Sverrisdottir OO, Daskalaki E, Skoglund P, Valdiosera CE, Carretero JM, Ferreras, JL et al. (2014) Un agricultor ibérico del Neolítico tardío exhibe afinidad genética con los agricultores escandinavos del Neolítico y un agricultor centroeuropeo de la Edad del Bronce. https://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-211155