
 El auge de la genómica antigua ha revolucionado nuestra comprensión de la prehistoria humana, pero este trabajo depende de la disponibilidad de muestras adecuadas. Presentaron un genoma humano antiguo completo secuenciado desde un trozo de resina de abedul masticada de 5700 años de antigüedad procedente de Dinamarca. Descubrieron que el individuo que masticó la resina era mujer y que estaba genéticamente más emparentada con los cazadores-recolectores occidentales de Europa continental que con los cazadores-recolectores de Escandinavia central. También que probablemente tenía piel oscura, pelo castaño oscuro y ojos azules. Los resultados destacan el potencial de la resina de abedul masticada como fuente de ADN antiguo.
El auge de la genómica antigua ha revolucionado nuestra comprensión de la prehistoria humana, pero este trabajo depende de la disponibilidad de muestras adecuadas. Presentaron un genoma humano antiguo completo secuenciado desde un trozo de resina de abedul masticada de 5700 años de antigüedad procedente de Dinamarca. Descubrieron que el individuo que masticó la resina era mujer y que estaba genéticamente más emparentada con los cazadores-recolectores occidentales de Europa continental que con los cazadores-recolectores de Escandinavia central. También que probablemente tenía piel oscura, pelo castaño oscuro y ojos azules. Los resultados destacan el potencial de la resina de abedul masticada como fuente de ADN antiguo.
Introducción
En esta publicación presentamos el trabajo de investigación de Theis Z.T. Jensen et all (ver en bibliografía). La brea de abedul es una sustancia de color negro-marrón que se obtiene calentando la corteza de abedul y se ha utilizado como adhesivo y agente de enmangue desde el Pleistoceno Medio. Pequeños trozos de este material orgánico se encuentran comúnmente en sitios arqueológicos en Escandinavia y más allá, y aunque su uso aún se debate a menudo muestran huellas de dientes, lo que indica que fueron masticados. La brea de abedul recién producida se endurece al enfriarse y se ha sugerido que masticarla era un medio para hacerla flexible de nuevo antes de usarla por ejemplo, para enmangar herramientas de piedra compuesta. También se han sugerido usos medicinales, ya que uno de los principales componentes de la brea de abedul, la betulina, tiene propiedades antisépticas. Esto está respaldado por una gran cantidad de evidencia etnográfica que sugiere que la brea de abedul se usaba como un antiséptico natural para prevenir y tratar dolencias dentales y otras afecciones médicas . Los ejemplos más antiguos de brea masticada encontrados en Europa datan del período Mesolítico y el análisis químico por cromatografía de gases-espectrometría de masas (GC-MS) ha demostrado que muchos de ellos estaban hechos de abedul ( Betula pendula ) .
Trabajos recientes de Kashuba et al han demostrado que piezas de brea de abedul masticada contienen ADN humano antiguo, que puede usarse para vincular la cultura material y la genética de poblaciones antiguas. En este estudio actual analizaron otra pieza de brea de abedul masticada que se descubrió en un yacimiento del Mesolítico tardío/Neolítico temprano en el sur de Dinamarca y demostraron que no solo contiene ADN humano antiguo sino también ADN microbiano que refleja el microbioma oral de la persona que masticó la brea, así como ADN vegetal y animal que puede haberse derivado de una comida reciente.
El ADN está tan excepcionalmente bien conservado que pudieron recuperar un genoma humano antiguo completo de la muestra (secuenciado a una profundidad promedio de cobertura de 2,3×), lo que es particularmente significativo ya que, hasta ahora, no se han recuperado restos humanos del yacimiento. Los resultados resaltan el potencial de la resina de abedul masticada como fuente de ADN humano y no humano antiguo, que puede utilizarse para arrojar luz sobre la historia de la población, el estado de salud e incluso las estrategias de subsistencia de las poblaciones antiguas.
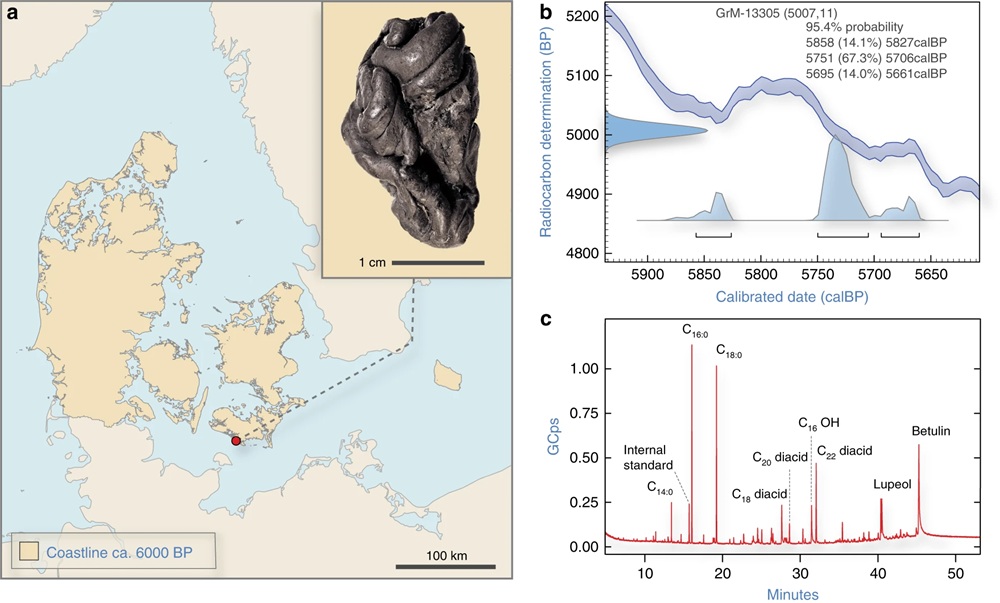
Un trozo masticado de brea de abedul del sur de Dinamarca. ( a ) Fotografía de la brea de abedul de Syltholm y su ubicación del hallazgo en el yacimiento de Syltholm en la isla de Lolland, Dinamarca (mapa creado usando datos de Astrup ). ( b ) Fecha calibrada para la brea de abedul de Syltholm (5858–5661 cal. BP; 5007 ± 7). ( c ) Cromatograma GC-MS de la brea de Syltholm que muestra la presencia de una serie de ácidos dicarboxílicos (diácido Cxx) y ácidos grasos saturados (Cxx:0) y metil 16-hidroxihexadecanoato (C OH) junto con los triterpenos betulina y lupeol, que son característicos de la brea de abedul .
Datación por radiocarbono y análisis químico
La datación por radiocarbono del espécimen arrojó una fecha directa de 5858–5661 cal. BP (GrM-13305; 5007 ± 11), que lo ubica al inicio del período Neolítico en Dinamarca. El análisis químico por espectroscopia infrarroja por transformada de Fourier (FTIR) produjo un espectro muy similar al de la brea de abedul moderna y la GC-MS reveló la presencia de los triterpenos betulina y lupeol, que son característicos de la brea de abedul. El espectro GC-MS también muestra un rango de ácidos dicarboxílicos y ácidos grasos saturados, que se consideran intrínsecos a la brea de abedul y, por lo tanto, respaldan su identificación.
secuenciación de ADN
Generaron aproximadamente 390 millones de lecturas de ADN para la muestra, casi un tercio de las cuales se pudieron mapear de forma única al genoma de referencia humano (hg19). Las lecturas humanas mostraron todas las características propias del ADN antiguo, incluyendo longitudes de fragmento promedio cortas, una mayor ocurrencia de
purinas antes de las roturas de cadena, y una mayor frecuencia de sustituciones aparentes de citosina (C) a timina (T) en los extremos 5' de los fragmentos de ADN y la
cantidad de contaminación humana moderna se estimó en alrededor del 1-3%. Además de las lecturas humanas, generaron alrededor de 7,3 Gb de datos de secuencia (68,8%) del paso antiguo que no se alinearon con el genoma de referencia humano.
Preservación del ADN y reconstrucción del genoma
Con más del 30%, el contenido de ADN endógeno humano en la muestra fue extremadamente alto y comparable al encontrado en dientes y huesos petrosos bien preservados . Usaron las lecturas humanas para reconstruir un genoma humano antiguo completo, secuenciado a una profundidad de cobertura efectiva de 2.3×, así como un genoma mitocondrial de alta cobertura (91×), que fue asignado al haplogrupo K1e . Para investigar más a fondo la preservación del ADN humano en la muestra, calcularon una tasa de desintegración molecular ( k , por sitio por año) y encontramos que es comparable a la de otros genomas humanos antiguos de regiones templadas.
Determinación sexual y rasgos fenotípicos
Con base en la proporción entre lecturas de alta calidad (MAPQ ≥ 30) que mapean a los cromosomas X e Y, respectivamente, determinaron el sexo del individuo cuyo genoma
recuperaron como femenino. Para predecir su color de cabello, ojos y piel, imputaron genotipos para 41 SNP incluidos en el sistema HIrisPlex-S y encontramos que probablemente tenía piel oscura, cabello castaño oscuro y ojos azules.

También examinaron el estado alélico de dos SNP vinculados con el haplotipo primario asociado con la persistencia de la lactasa en humanos y descubrimos que portaba el alelo ancestral para ambos, lo que indica que no era persistente a la lactasa.
Afinidades genéticas
Llamamos a 593.102 polimorfismos de un solo nucleótido (SNP) en nuestro genoma antiguo que previamente habían sido genotipados en un conjunto de datos de >1000 individuos actuales de un conjunto diverso de poblaciones euroasiáticas, así como >100 genomas antiguos publicados previamente. La muestra se agrupa con cazadores-recolectores occidentales (WHG). Las estimaciones de compartición de alelos basadas en las estadísticas f muestran la misma afinidad general con los WHG. Esto también se refleja en el análisis qpAdm que demuestra que un modelo unidireccional simple que asume una ascendencia 100% WHG no puede rechazarse a favor de modelos más complejos. Para probar formalmente este resultado, calcularon dos conjuntos de estadísticas D de la forma D (Yoruba, EHG/Barcın; prueba, WHG) y no encontraron evidencia de niveles significativos de EHG o flujo genético de agricultores neolíticos.
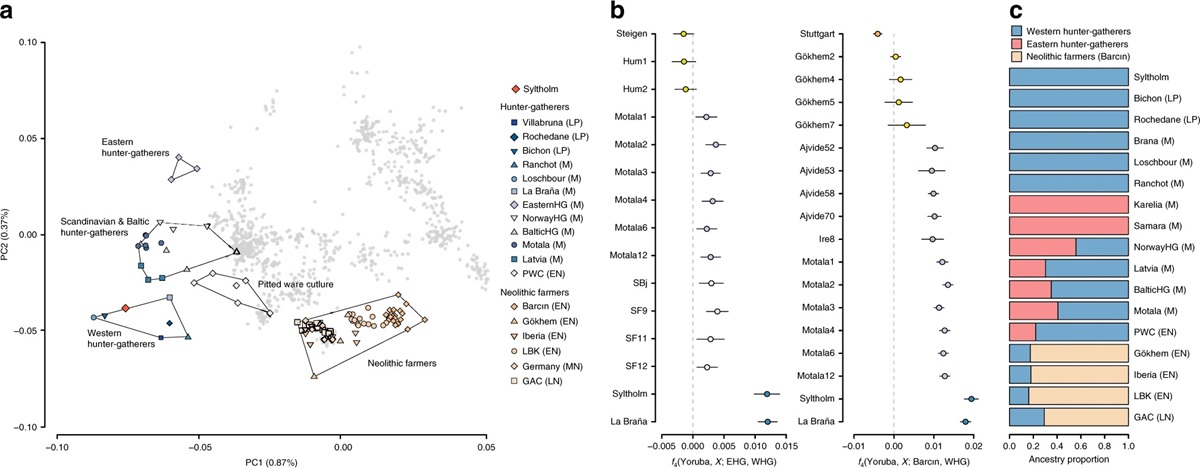
Afinidades genéticas del individuo Syltholm. a Análisis de componentes principales de individuos euroasiáticos modernos (en gris) y una selección de más de 100 genomas antiguos publicados previamente, incluyendo el genoma de Syltholm. Los individuos antiguos fueron proyectados sobre la variación moderna (ver Métodos). b Estimaciones de compartición de alelos entre el individuo Syltholm, otros individuos mesolíticos y neolíticos, y WHG versus EHG y agricultores neolíticos, respectivamente, medidos por el estadístico f (Yoruba, X ; EHG/Barcın,WHG). c Proporciones de ascendencia basadas en qpAdm, especificando WHG, EHG y agricultores neolíticos (Barcın) como posibles poblaciones de fuentes ancestrales. Cultura de cerámica con hoyos PWC , LBK Cerámica de banda lineal, GAC Cultura de ánforas globulares, LP Paleolítico tardío, M Mesolítico, EN Neolítico temprano, MN Neolítico
medio, LN Neolítico tardío.
Resumen
Extrajeron y secuenciaron con éxito ADN antiguo de un trozo de brea de abedul masticada de 5700 años de antigüedad del sur de Dinamarca. Además de un genoma humano antiguo completo (2,3×) y mitogenoma (91×), recuperamos ADN vegetal y animal, así como ADN microbiano de varios taxones orales. El análisis de las lecturas humanas reveló que el individuo cuyo genoma recuperamos era femenino y que probablemente tenía piel oscura, cabello castaño oscuro y ojos azules. Esta combinación de rasgos físicos se ha observado previamente en otros cazadores-recolectores europeos, lo que sugiere que este fenotipo estaba muy extendido en la Europa mesolítica y que la propagación adaptativa de la pigmentación clara de la piel en las poblaciones europeas solo ocurrió más tarde en la prehistoria. También tenía los alelos asociados con la no persistencia de la lactasa, lo que encaja con la noción de que la persistencia de la lactasa en adultos sólo evolucionó bastante recientemente en Europa, después de la introducción de la producción lechera con la revolución neolítica .
Desde el punto de vista de la genética de poblaciones, el genoma humano también ofrece nuevas perspectivas sobre el poblamiento temprano del sur de Escandinavia. Estudios recientes de genomas de antiguos cazadores-recolectores de Suecia y Noruega han demostrado que, tras el retroceso de las capas de hielo hace unos 12-11 ka años, Escandinavia fue colonizada por dos rutas separadas, una desde el sur (presumiblemente a través de Dinamarca) y otra desde el noreste, a lo largo de la costa de la actual Noruega. Esto se ve respaldado por el hecho de que los cazadores-recolectores de Escandinavia central tienen diferentes niveles de ascendencia WHG (cazadores-recolectores del oeste) y EHG (cazadores-recolectores del este), que llegaron a Escandinavia central desde el sur y el noreste, respectivamente. Aunque solo analizaron un solo genoma, el hecho de que el individuo Syltholm no tenga ninguna ascendencia EHG confirma este escenario y sugiere que los EHG no llegaron al sur de Dinamarca en este punto de la prehistoria.
El genoma de Syltholm (5700 años cal. BP) data del período inmediatamente posterior a la transición Mesolítico-Neolítico en Dinamarca. Culturalmente, este período está marcado por la transición de la cultura Ertebølle del Mesolítico Tardío (c. 7300–5900 cal. BP) con sus artefactos de piedra lascada y sus típicas hachas de asta en forma de T, a la cultura del vaso de embudo del Neolítico temprano (c. 5900–5300 cal. BP) con su cerámica característica, artefactos de sílex pulido y plantas y animales domesticados. En Dinamarca, la transición de la caza y la recolección a la agricultura a menudo se ha descrito como un proceso relativamente rápido, con cambios drásticos en los patrones de asentamiento y las estrategias de subsistencia. Sin embargo, aún no está claro en qué medida esta transición fue impulsada por la llegada de comunidades agrícolas en oposición a la adaptación local de las prácticas agrícolas por parte de las poblaciones residentes de cazadores-recolectores.
Estos análisis han demostrado que el individuo Syltholm no tiene ascendencia de agricultores neolíticos, lo que sugiere que el impacto genético de las comunidades agrícolas neolíticas en el sur de Escandinavia podría no haber sido tan instantáneo o generalizado como se creía. Si bien este ADNmt recuperado pertenece al haplogrupo K1e, que se asocia más comúnmente con las comunidades agrícolas tempranas, existe evidencia creciente que sugiere que este linaje ya estaba presente en la Europa mesolítica. En general, la falta de ascendencia de agricultores neolíticos es consistente con la evidencia de otras partes de Europa, lo que sugiere que los grupos de cazadores-recolectores genéticamente distintos sobrevivieron durante mucho más tiempo de lo que se suponía anteriormente. Estos "sobrevivientes" WHG podrían haber desencadenado el resurgimiento de la ascendencia de cazadores-recolectores que se propone que ocurrió en Europa central entre 7000 y 5000 BP.
Bibliografia:
https://doi.org/10.1038/s41467-019-13549-9