
 El yacimiento del Alto de la Cruz de Cortes de Navarra se descubrió casualmente en 1947, se trataba de un yacimiento de gran interés para el conocimiento de la implantación de la metalurgia del hierro en el Occidente europeo. La transición del Bronce Final a la Edad del Hierro en la península Ibérica supuso un cambio no generalizado en las costumbres mortuorias, que pasaron de la inhumación principal a la cremación de los difuntos. Todos serán incinerados menos los neonatos.
El yacimiento del Alto de la Cruz de Cortes de Navarra se descubrió casualmente en 1947, se trataba de un yacimiento de gran interés para el conocimiento de la implantación de la metalurgia del hierro en el Occidente europeo. La transición del Bronce Final a la Edad del Hierro en la península Ibérica supuso un cambio no generalizado en las costumbres mortuorias, que pasaron de la inhumación principal a la cremación de los difuntos. Todos serán incinerados menos los neonatos.
Se asocia este cambio de costumbre a la entrada de los “campos de urnas ” sobre el siglo X a.C. y cuya costumbre remontó el río Ebro hasta llegar casi hasta Navarra. Actualmente casi nadie sostiene que los grupos de los campos de urnas fueran un ente cultural homogéneo; la opinión generalizada es que se trató simplemente de una moda que se extendió por Europa debido a préstamos culturales o, en ciertos casos, a pequeños movimientos de pueblos diferenciados entre sí. (https://es.wikipedia.org/wiki/Cultura_de_los_campos_de_urnas)

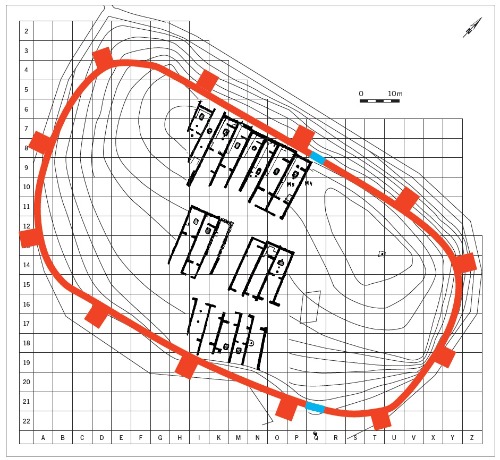
(Recreación por Landa, de enterramiento intramuros)
La cremación dificulta los análisis moleculares (análisis isotópicos, ADN antiguo) de los habitantes antiguos. Pero gracias a otro singular rito, donde niños pequeños y a menudo recién nacidos, estaban exentos del ritual de cremación, tenemos acceso a las inhumaciones intramuros en el interior de los edificios ibéricos.

Es una suerte poder ver el estudio los genomas de 37 niños enterrados intramuros encontrados en tres asentamientos navarros de la Edad del Hierro Temprana, datados entre c. 800 y 450 a. C. Los análisis genéticos de estos individuos recientemente estudiados por Luka Papac, Patxuka de Miguel Ibáñez y otros, amplían la comprensión de la antigua Iberia al revelar su diversidad genética.
Estos niños navarros de la edad de Hierro muestran una menor influencia de la ascendencia mediterránea comparándolos con los individuos de la edad del Hierro publicados anteriormente del norte de España.
Si vemos la figura 16 (el mapa A es la situación actual euroasiática), veremos en el mapa B que los niños del Alto de la Cruz (IA Navarra en Rojo) están situados en la misma zona PC2 (parámetro que está correlacionada con ascendencia Caucásica y Oriente Próximo) de los actuales españoles del norte y franceses del sur (con un índice PC2 aprox. -0.015).
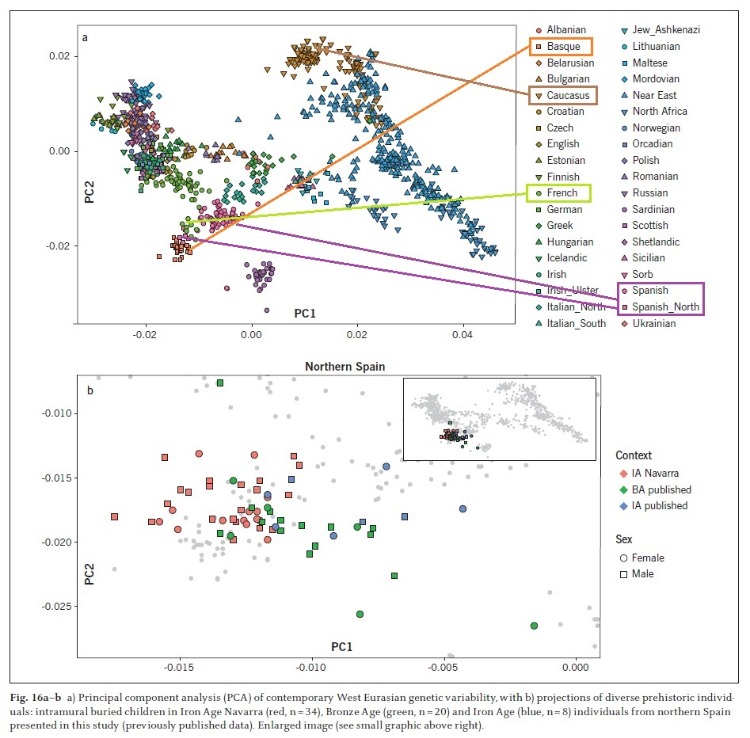
Fig. 16:
a) Análisis de componentes principales (PCA) de la variabilidad genética contemporánea de Eurasia occidental.
b) Proyecciones de diversos individuos prehistóricos: niños enterrados intramuros en Navarra de la Edad del Hierro (rojo), individuos de la Edad del Bronce (verde) y de la Edad del Hierro (azul) del norte de España(datos publicados previamente). Imagen ampliada (ver gráfico pequeño arriba a la derecha).
En la misma imagen o mapa genético (el parámetro PC1 que se correlaciona con ascendencia del Mediterránea) se puede ver como los individuos del norte ibérico de la edad del Bronce y los anteriormente publicados de la edad del Hierro tenían una mayor influencia del sur mediterráneo. Los datos genéticos de estos individuos llenan un vacío importante en el registro arqueo genético del norte de España, aun a estudiar.
Poco acertado parece relacionar ascendencia Esteparia con ascendencia Caucásica u Oriente Próximo, cuna de la agricultura. Para muestra, abajo tenemos un mapa de la ascendencia esteparia en Europa, incluida nuestra zona ibérica y la Navarra en cuestión.
HAPLOGRUPOS MITOCONDRIALES femeninos en Alto de La Cruz.
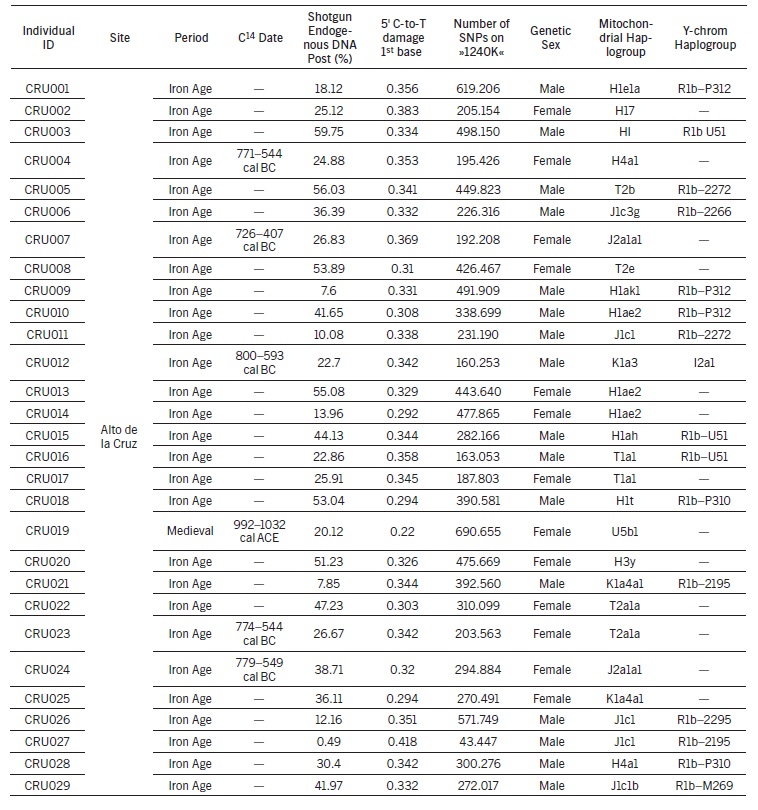
Del total de 37 niños de la Edad del Hierro estudiados por Luka Papac y Patxuka, 29 lo fueron muestreados en Alto de la Cruz (CRU), 6 en Las Eretas (ERE) y 2 niños en Castejón de Bargota (BGO) en la región de Navarra de la Primera Edad del Hierro (800-40 a. C.). De todos ellos, en Alto de la Cruz fueron encontrados los diferentes haplogrupos mitocondriales:
1 individuos tipo U5
5 individuos tipo J1c
2 individuos tipo J2a
3 individuos tipo K1a4a1
2 individuos tipo T1a
4 individuos tipo T2a
8 individuos tipo H1 de un total de 12 tipo H.
El DNA-mt tipo J.
Otro haplogrupo mitocondrial antiguo y típico de Eurasia Occidental es el JT, muy común en los antiguos etruscos. Está compuesto a su vez por los haplogrupos J y T.
El tipo J tiene un probable origen en el Medio Oriente hace 30.000 y 40.000 años. Este haplogrupo sobrevivió a la era glacial del Pleistoceno, cuyos individuos se protegieron de las condiciones climáticas en la región conocida como franco-cantábrica, al norte de España y sureste de Francia, y después se asentó en Europa con la llegada de la agricultura. Las frecuencias ACTUALES más importantes de J1c son en Islandia y de J2a en el Sur de Europa.
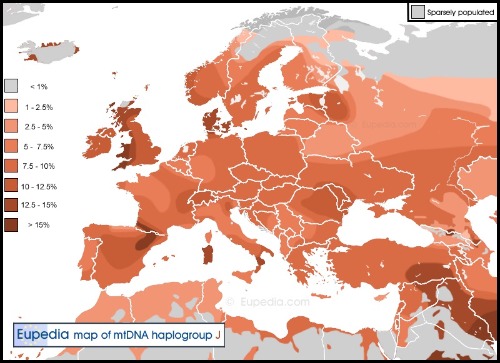
El J1c es tan raro en Oriente Medio hoy en día, que se puede imaginar que fuera un linaje de cazadores-recolectores que habitaron el Mediterráneo oriental y el sur de los Balcanes durante el Epipaleolítico, pero que su difusión por el resto de Europa no tuvo lugar hasta el período Neolítico. Lazaridis et al.(2016) analizaron 44 muestras del antiguo Cercano Oriente, incluidos agricultores neolíticos de Jordania e Irán occidental, así como muestras del Calcolítico y la Edad del Bronce de Armenia y el Levante, pero no encontraron ningún J1c. Sobre las muestras neolíticas de Europa Central, se puede asumir que los linajes maternos J1c y J2b1a NO FUERON TRAÍDOS por los recién llegados en las invasiones indoeuropeas durante la Edad del Bronce.
El DNA-mt tipo T1.
Del T1 las frecuencias ACTUALES más importantes son en Europa y Egipto. El T2 está ampliamente disperso desde Irlanda, Portugal, Marruecos, hasta la India, Kazajistán y pueblos túrquicos y mongoles de Siberia. Es mayoritario en samaritanos con 56%. El haplogrupo T está compuesto por dos ramas principales, T1 y T2. Ambas tienen distribuciones muy diferentes, diametralmente opuestas en la mayoría de las regiones.
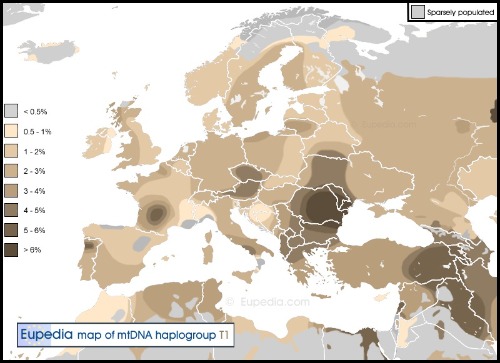
El haplogrupo T1 no se encuentra entre los saami, los judíos o los ávaros del Cáucaso, y es extremadamente raro en Jordania, Marruecos, el norte de España, Bosnia y Croacia. Los haplogrupos T* (quizás T1a) y T2b se han encontrado en esqueletos de cazadores-recolectores del Mesolítico tardío de Rusia y Suecia respectivamente.
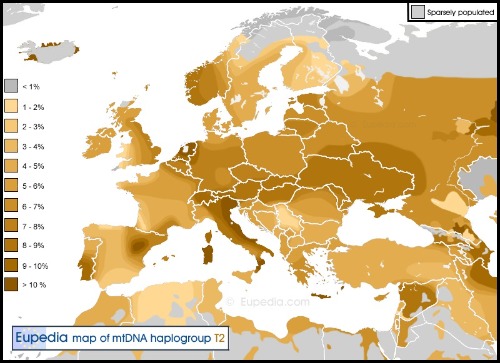
Es la mejor evidencia hasta el momento de que el haplogrupo T estaba presente en Europa antes de que el continente fuera recolonizado por agricultores neolíticos. Sin embargo, dado que las muestras son contemporáneas a las culturas neolíticas del resto de Europa, no es seguro que los linajes T no surgieran a través de matrimonios mixtos entre agricultores y cazadores-recolectores.
El DNA-mt tipo K1a.
El haplogrupo K, aunque parece haber estado ausente entre los cazadores-recolectores occidentales (WHG) que ocuparon Europa occidental y central antes del Neolítico, esta en Lola (WHG de Dinamarca). Los subclados K1a, K1b y K2a se encontraron entre los agricultores del Neolítico temprano (ENF) del Cercano Oriente y, posteriormente, entre los agricultores europeos tempranos (EEF). Como hemos visto el grupo K es un subclado del haplogrupo U8b creado en algún momento entre 20.000 y 38.000 años atrás. El K1a es el subclado más común y diverso en la Europa actual, y ya era, con diferencia, el subclado más común entre los agricultores neolíticos. La gran mayoría de las muestras neolíticas de Europa central y occidental eran K1a.
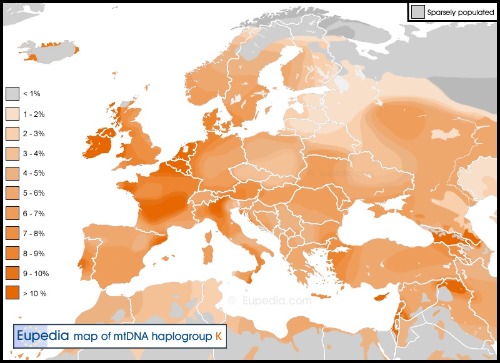
Este K1a del Alto de la Cruz en Navarra es K1a4a1 (común en Cataluña y Alemania). El K1a4a1 es el subclado K más común en Europa occidental, Alemania y Escandinavia en la actualidad. El K1a4 probablemente fue uno de los principales subclados K1a entre los agricultores neolíticos.
El DNA-mt tipo H.
El haplogrupo H es el linaje materno mayoritario en el Alto de la cruz y el más común y más diverso en Europa, en la mayor parte del Cercano Oriente o en la región del Cáucaso, y posee aproximadamente 90 subclados basales identificados hasta la fecha. La mutación que define al haplogrupo H se produjo hace al menos 25.000años, y quizá cerca de 30.000 años. Se desconoce su lugar de origen, pero probablemente se encontraba en algún lugar del noreste del Mediterráneo (Balcanes, Anatolia o Levante), posiblemente incluso en Italia.
En un yacimiento de la cultura de la cerámica Cardial se recuperaron 17 secuencias de ADNmt, incluidas tres que pertenecían al haplogrupo H1 y tres al H3. Los hombres pertenecían al haplogrupo G2a de ADN-Y del Cercano Oriente y al haplogrupo I2a del Mesolítico europeo. Sin embargo, es digno de mención que los hombres I2a pertenecían exclusivamente a H1 y H3, una evidencia potente, aunque no concluyente, de que H1y H3 eran linajes que pertenecían a cazadores-recolectores mesolíticos del suroeste de Europa.
Las distribuciones modernas de H1 y H3 respaldan una recolonización tardo glacial de Europa occidental y central desde el refugio franco-cantábrico.
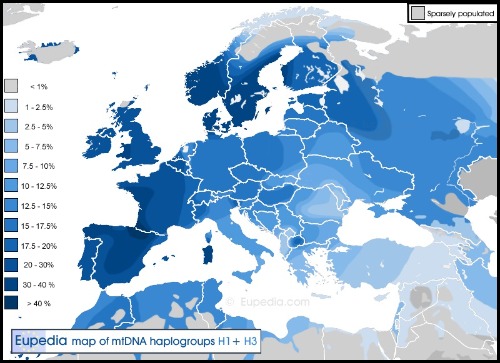
El individuo CRU004 y el CRO028 encontrado en Navarra del haplogrupos H4 también se encontraría entre los europeos del Mesolítico o del Paleolítico Superior Tardío basándose en su presencia exclusiva en Europa hoy. Este H4 lo vemos en Navarra neolítica y se encuentra hoy en día tanto entre los vascos como entre los sardos, dos poblaciones con un alto porcentaje de ascendencia europea mixta mesolítica y neolítica. Sin embargo, H4nunca se encontró entre los primeros agricultores neolíticos del Cercano Oriente o el Sureste de Europa.
HAPLOGRUPOS Y masculinos en el Alto de La Cruz
El DNA-Y tipo R-L51
R-L51 es típico de Europa occidental, es el verdadero haplotipo representativo del Atlántico y se habría originado en Europa central hace unos 6.000 años, aunque hay otra rama paralela entre los yamna. El R1b-L51es un clado intermedio entre los subgrupos L23 oriental y L11+ (P310+) occidental de R1b. Se encuentra en frecuencias extremadamente bajas desde Irán hasta Inglaterra, con una incidencia relativamente mayor en Europa central. Aún no está claro si surgió en Europa o en Anatolia, pero vale la pena mencionar que los subclados principales R1b-P312 (U152, M167, L21) tienen su mayor variación en Francia, por lo tanto, se cree que L51 se formó en el sur de Francia (y no en Europa del este) durante el Neolítico tardío a partir de personas R1b L23 que habían llegado de Italia.
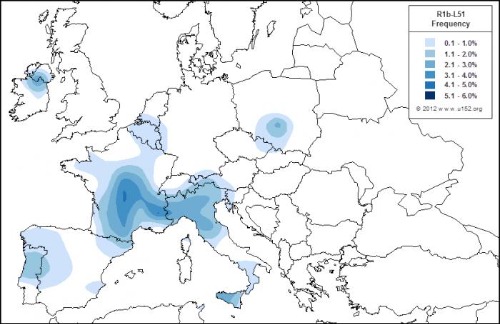
(mapa de R1b-L51 durante el neolítico; fuente Europedia)
Este mapa muestra altas frecuencias de este subclado en el sur/centro de Francia, así como en el norte de Italia. R1b L51 parece estar ausente de los Balcanes y Europa del este en general (excepto el sur de Polonia). Apuntar que L51 y su sucesor L11 se extendieron desde Europa occidental a Europa central y no al revés, lo que los hace no relacionados con las culturas de Europa central de la Edad del Bronce (Unetice, Urnfields, etc.).
Nuestro árbol filogenético del haplogrupo R1b muestra claramente que, además de la conocida rama R-L51 predominante en Europa occidental, existe otra rama R-GG400 distinta en Europa oriental. Los marcadores que definen esta rama oriental se revelaron en un conjunto de secuencias del cromosoma Y publicadas (Karmin et al. 2015; Hal-last et al. 2015; Batini et al. 2015).
La rama oriental R-GG400 es de igual nivel filogenético y edad similar a la rama R-L51, pero con una distribución geográfica contrastante. La distribución de la rama occidental R-L51 es bien conocida, se encuentra en frecuencias más bajas en Europa del Este (Myres et al. 2011). Los datos recopilados sobre la distribución geográfica de la rama oriental de R-GG400 son escasos, pero ubican su área principal en las estepas de Europa del Este y Asia Occidental (Karmin et al. 2015) y muy escasa o nula presencia en el occidente europeo.
Las antiguas muestras Yamnaya se encuentran en la rama R-GG400 "oriental" del haplogrupo R1b-L23, lo que demuestra que los descendientes paternos de los Yamnaya todavía viven en la estepa póntica y que "la antigua población Yamnaya no fue una fuente importante de linajes paternos en los actuales europeos occidentales", según Balanovsky, O.; Chukhryaeva, M.; Zaporozhchenko, V. del Instituto de Genética General Vavilov, Moscú, Rusia y del Centro de Investigación en Genética Médica, Moscú, Rusia. (2017, "LADIFERENCIACIÓN GENÉTICA ENTRE POBLACIONES DE TIERRAS ALTAS Y BAJAS DA FORMA ALPAISAJE CROMOSÓMICO Y DE ASIA OCCIDENTAL"(https://labs.icb.ufmg.br/lbem/pdf/Balanovsky2017HGlowlandAsia.pdf)
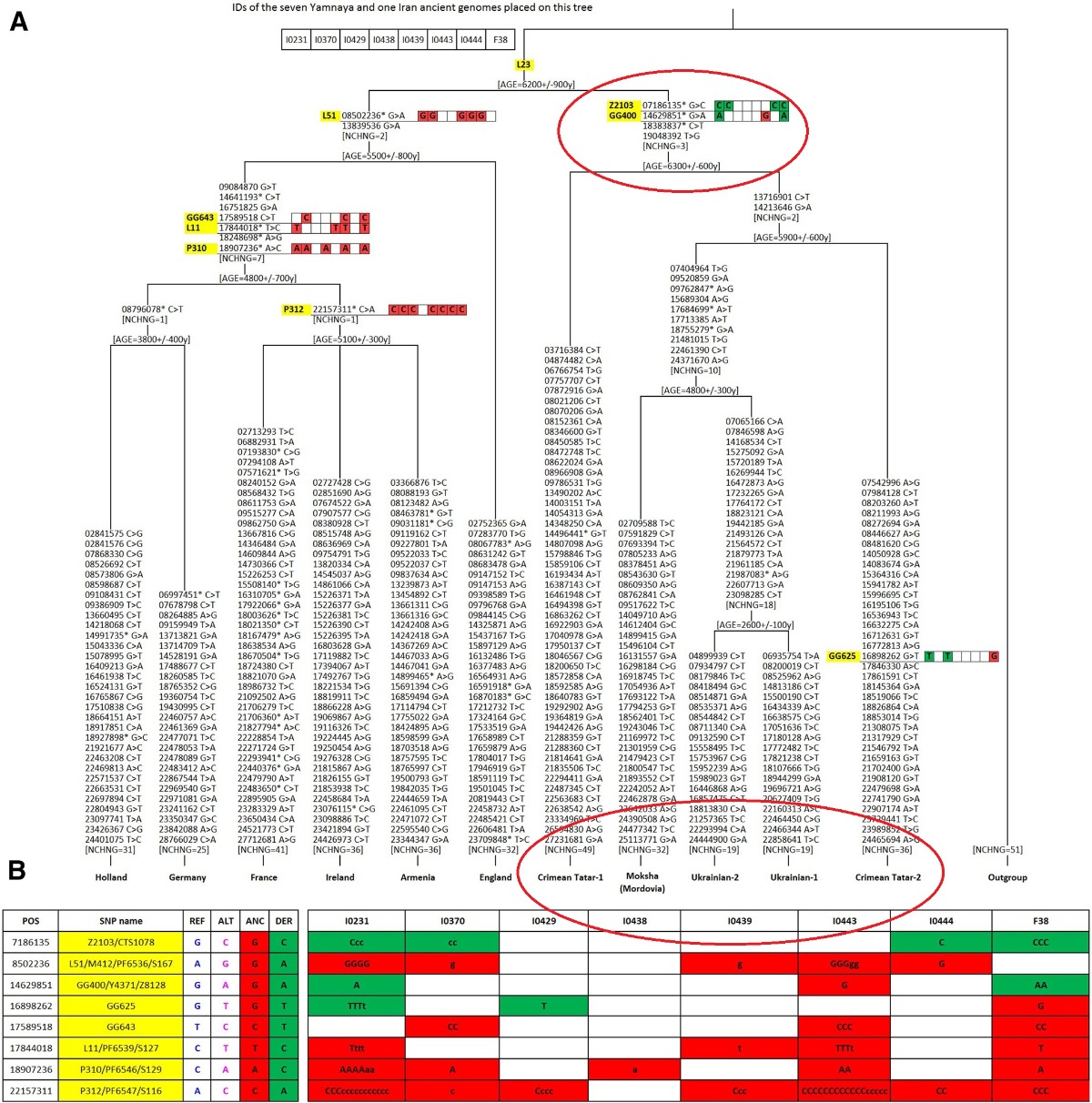
(PINCHA IMAGEN para agrandar )
El DNA-Y tipo R-P310
El R-P310 se formó cuando se separó del antepasado R-L51 y se crea alrededor 4.100 a.C. Es el antepasado de R-L151, R-FT377377, R-FT186340, & R-FT123498.
El DNA-Y tipo R-P312
El R-P312 (P312/S116). S116 incluye subclados asociados con lenguas no indoeuropeas como el vasco y las antiguas lenguas gasconas e ibéricas. Típico de Europa occidental. Se halló en España en restos de hace 3.300 a.C. y se formó cuando se separó de su antepasado R-L151. Sus linajes, son la mayoría de los R1b europeos menos los greco-etruscos R1b-L23, los germánicos R1b-U106 y R1b-L238, y los proto-celto-germánicos L11, L51y L150. Es el antepasado de al menos 9 linajes descendientes conocidos como R-DF27; Z272 (>M153 vascos); S228/Z198; R-PF6570/S28/U152; R-DF19 (germánicos, sajones); R-L238 (nórdicos); R-A9063; R-Y18211; S461/Z290; R-BY25512; R-Z29913; R-Z46516.
R1b-P312 es el subgrupo occidental de L11, el más común en las áreas de habla celta y romance occidental. Tiene varios subclados que tienen un patrón geográfico claro, siendo los más grandes U152 y L21. En su formaP312* (U152-L21-), es más común en Iberia (35-50%) y Francia (20-35%), pero también ocurre con frecuencia en Irlanda, Inglaterra y Suiza (10-20%). Es menos frecuente (5-10%) en el norte de Italia, Alemania, Países Bajos y Escandinavia. Su pequeño subgrupo M153 solo se encuentra entre los vascos. Su edad estimada es similar a la de U106, aproximadamente 5500-4000 años antes del presente. La diversidad apunta a Francia como el probable lugar de origen, y generalmente está relacionado con la expansión de las poblaciones de habla celto-itálica. Sin embargo, puede representar un pueblo más antiguo, el relacionado con los vascos.
El DNA-Y tipo R1b-M269
Tiene una antigüedad superior a 8.000 años, y por ello pertenecen al Mesolítico, por lo que son anteriores a la entrada en Europa de los primeros agricultores, y lógicamente a la expansión indoeuropea. Los restos de Villabruna (Italia), con 14.000 años de antigüedad porta el haplogrupoR1b, que por el genotipo que muestra de OJOS AZULES, podría estar emparentado o incluso ser antecesor deR1b-M269. Según la genetista Patricia Balaresque y otros, aunque estudios anteriores sugirieron un origen paleolítico, demostraron que R1b-M269 coincide con la llegada y expansión de la agricultura en Europa (Balaresque P, Bowden GR, Adams SM, Leung HY, King TE, Rosser ZH, et al. 2010. Un origen predominantemente neolítico para los linajes paternos europeos).

Este es otro marcador al que asocian “alegremente” un origen entre los Yamnaya, pero George BJ Busby de la University of Oxford, UK confirmó que la distribución espacial del haplogrupo M269 puede dividirse (mediante R-S127) en linajes europeos y euroasiáticos occidentales:
<< En el futuro las comparaciones de secuencias proporcionarán fechas sólidas para este haplogrupo. Por ahora, no podemos ofrecer ninguna fecha sobre la edad de R-M269 o RS127, pero creemos que nuestros análisis STR sugieren que las estimaciones de edad recientes de R-M269 y R-S116 probablemente sean más jóvenes que los valores verdaderos, y la homogeneidad de la variación de STR y la distribución de los subtipos en todo el continente SON INCONSISTENTES con la hipótesis de la difusión neolítica del linaje del cromosoma Y R-M269.>>
Resulta curioso que el individuo con haplogrupo R1b-M269 más antiguo tenga 8.716 años (del Mesolítico temprano y código ref. UZZ026) y aparezca en el occidente europeo, en SICILIA (Italia) y no en Asia, además de que tenía OJOS AZULES y piel morena. En Inglaterra hay un individuo R1b-M269 de 5.700 años (Ingleborough Hill, Fox Holes Cave) y otro de 4.900 años (Blaydon, Tyne and Wear, Summerhill) que se asocian a la cultura del vaso campaniforme. Otro individuo R1b-M269 con 8.331 años y código I3883 apareció en Shanidar (Irak). Apareció en Jordania un individuo M269 con 7.950 años y código I8554. En Letonia, con 7.268 años está el individuo Cazador-Recolector con código I4630, portaba marcador M269 (DNAmt U5a2c). En Samara, Rusia, está el individuo Cazador-Recolector con código I0124 y 5.594 años de M269 (DNAmt U5a1d), con piel pálida, cabello marrón, ojos azules.
La región integrada por Ucrania, Rumanía y Moldavia presentan R1b en el periodo antiguo con frecuencias muy escasos con valores por debajo de 0’2 a lo largo del tiempo excepto hace 2.999-2.000 años, cuando toma un valor máximo de 0’5. En Ucrania aparecen los primeros individuos con el haplogrupo R1b-M269 hace 4.700 años.
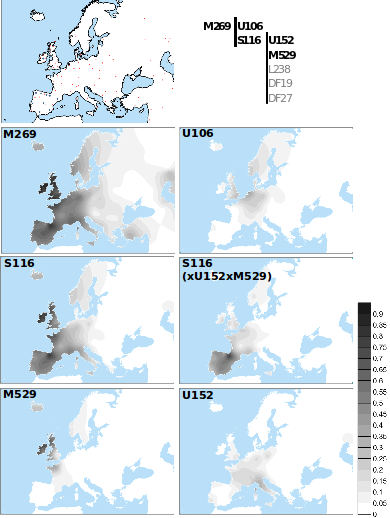
La presencia del R1b-M269 en occidente no cuadra, ni con la expansión Yamnaya, ni con su origen asiático. Si bien el R1b está presente en los kurganes más antiguos (4.000 - 2.000 a.C), hoy resulta casi ausente en la región que se presume es la patria de origen de las poblaciones indoeuropeas (Ucrania y las estepas euroasiáticas) cuya población es principalmente R1a. Sabemos que las primeras elites enterradas en los kurganes eran principalmente de Y-ADN tipo I2a; R1a; R1b-L23; R1b-M269 y sobre todo R1b-Z2103, muestras relativamente jóvenes si las comparamos con el de Sicilia de 8.700 años o Ingleborough Hill de 5.700 años.
Es muy posible que el origen desde la variante M-269 de R1b sea la rama occidental del hombre de Villabruna. Un varón cazador recolector de piel morena y ojos azules enterrado en Italia perteneciente a la cultura epi-gravetiense extendida por gran parte de Europa occidental llegando hasta la zona ucraniana que después ocuparon los pastores nómadas Yamnaya.
Laura Valverde et al. del Grupo de Investigación BIOMICs, Centro de Investigación Lascaray, Universidad del País Vasco UPV/EHU, en su publicación de “New clues to the evolutionary history of the main European paternal lineage M269: dissection of the Y-SNP S116 in Atlantic Europe and Iberia”, llegaron a la misma conclusion que Busby y afirman:
<< es poco probable que una llegada a Europa de M269 durante el Neolítico haya generado un escenario tan complejo de expansiones para sus sublinajes>> / <<El escenario propuesto aquí sería más compatible con una llegada de M269 desde el Este en tiempos del Paleolítico>>
Valverde propone que los linajes R que evolucionaron en el ESTE desde tiempos paleolíticos, podrían haberse expandido hacia el Oeste DURANTE EL NEOLÍTICO, mezclándose así con otros linajes R que llegaron a Europa Occidental EN TIEMPOS PALEOLÍTICOS y que evolucionaron de forma independiente en estos territorios occidentales. Valverde destaca:
<<Los movimientos post-Neolíticos pueden estar enmascarando y confundiendo los rastros más antiguos>>
Bibliografia:
- INTRAMURAL CHILD BURIALS IN IRON AGE NAVARRA: HOW ANCIENT DNA CAN CONTRIBUTE TO HOUSEHOLD ARCHAEOLOGY. Luka Papac, Patxuka de Miguel Ibáñez, Adam B. Rohrlach, Javier Armendáriz, Marcello Peres,Thiseas C. Lamnidis, Angela Mötsch, Stephan Schiffels, and Roberto Risch
- The peopling of Europe and the cautionary tale of Y chromosome lineage R-M269. George B. J. Busby et al. https://www.researchgate.net/publication/51593643_The_peopling_of_Europe_and_the_cautionary_tale_of_Y_chromosome_lineage_R-M269
- New clues to the evolutionary history of the main European paternal lineage M269: dissection of the Y-SNP S116 in Atlantic Europe and Iberia. Laura Valverde et al. del Grupo de Investigación BIOMICs, Centro de Investigación Lascaray, Universidad del País Vasco UPV/EHU, https://www.nature.com/articles/ejhg2015114