
 Este es un resumen de un estudio de 2023 sobre paleogenomica que comprende desde del Paleolítico Superior hasta el Neolítico realizado por científicos del Instituto de Ciencias Arqueológicas, Departamento de Geociencias, el Centro Senckenberg para la Evolución Humana y el Paleoambiente de la Universidad de Tubinga y el Departamento de Arqueogenética, Instituto Max Planck de Antropología Evolutiva de Leipzig (Alemania).
Este es un resumen de un estudio de 2023 sobre paleogenomica que comprende desde del Paleolítico Superior hasta el Neolítico realizado por científicos del Instituto de Ciencias Arqueológicas, Departamento de Geociencias, el Centro Senckenberg para la Evolución Humana y el Paleoambiente de la Universidad de Tubinga y el Departamento de Arqueogenética, Instituto Max Planck de Antropología Evolutiva de Leipzig (Alemania).
Posth, C., Yu, H., Ghalichi, A. et al. Paleogenómica de los cazadores-recolectores europeos del Paleolítico Superior al Neolítico. Nature 615 , 117–126 (2023). https://doi.org/10.1038/s41586-023-05726-0 https://www.nature.com/articles/s41586-023-05726-0
El conocimiento de la relación genética y la estructura de los antiguos cazadores recolectores es limitado, debido a la escasez y mala conservación molecular de los restos humanos de ese período.
Cosimo Posth, He Yu et all. analizaron 356 genomas de antiguos cazadores-recolectores, incluidos 116 individuos nuevos de 14 países de Eurasia occidental y central, que abarcan entre 35.000 y 5.000 años atrás.
Identificaron un perfil de ascendencia genética en individuos asociados a conjuntos Gravetienses del Paleolítico Superior de Europa occidental que es distinto de los grupos contemporáneos relacionados con esta cultura arqueológica en Europa central y meridional , pero se asemeja al de los individuos precedentes asociados con la cultura Auriñaciense.
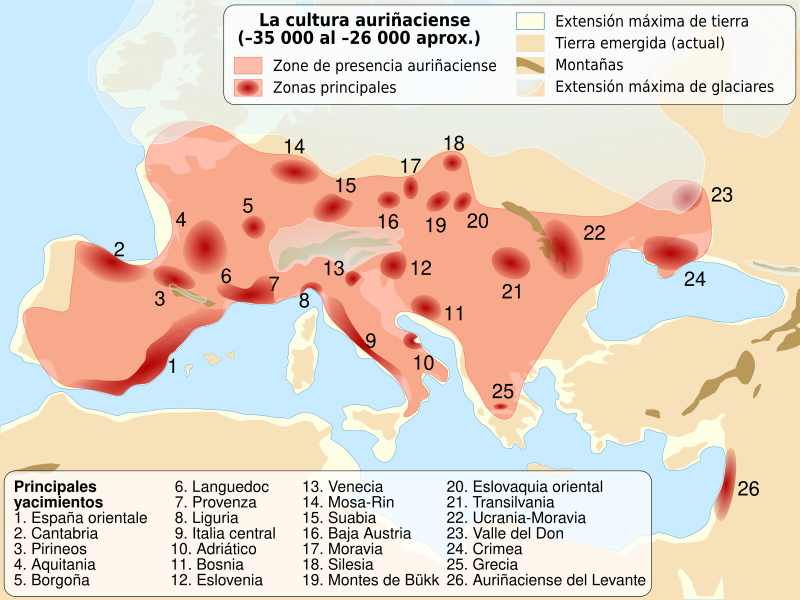
Este perfil de ascendencia sobrevivió durante el Último Máximo Glacial (hace 25.000 a 19.000 años) en poblaciones humanas del suroeste de Europa asociadas con la cultura Solutrense, y con la posterior cultura Magdaleniense que se re-expandió hacia el noreste después del Último Máximo Glacial.
Revelaron una renovación genética en el sur de Europa que sugiere un reemplazo local de grupos humanos alrededor de la época del Último Máximo Glacial, acompañado de una dispersión de norte a sur de poblaciones asociadas con la cultura Epigravetiense.
Desde hace al menos 14.000 años, una ascendencia relacionada con esta cultura se extendió desde el sur por el resto de Europa, reemplazando en gran medida el acervo genético asociado al Magdaleniense. Después de un período de mezcla limitada que abarcó el comienzo del Mesolítico, encontramos interacciones genéticas entre cazadores-recolectores de Europa occidental y oriental, que también se caracterizaron por marcadas diferencias en variantes fenotípicamente relevantes.
Antes del LGM
La cultura gravetiense fue una de las culturas del Paleolítico Superior más ampliamente distribuidas en Eurasia occidental antes del UMG y se considera un mosaico cultural con variaciones regionales. Sus genomas se originan en Europa central y meridional, dejando sin describir el perfil genético de los grupos humanos asociados al gravetiense de Europa occidental y suroccidental.
Para trazar una matriz y hacer estadísticas tienen en cuenta la presencia de tres grupos distintos:
- un grupo anterior a 40 ka con individuos de los yacimientos de Ust'Ishim, Bacho Kiro, Zlatý kůň y Peştera cu Oase
- un grupo de Věstonice que incluye individuos asociados al Gravetiense de yacimientos del centro-este y sur de Europa (Dolní Věstonice, Pavlov, Krems-Wachtberg, Paglicci y Ostuni).
- un grupo de Fournol que comprende individuos asociados al Gravetiense de yacimientos de Europa occidental y suroccidental (Ormesson, La Rochette, Fournol y dos yacimientos de cuevas de Serinyà (Mollet III y Reclau Viver)).
El grupo de Věstonice descrito anteriormente, incluye un nuevo individuo de 29.000 años de la cueva de Paglicci en el sur de Italia, está estrechamente relacionado con los genomas previamente publicados de Sunghir y Kostenki en el oeste de Rusia, que datan de 34 ka y 32 ka, respectivamente. El grupo de Fournol está estrechamente relacionado con los individuos asociados al Auriñaciense de Bélgica que datan de 35 ka (Goyet Q116-1 y Goyet Q376-3). Otra población asociada al Gravetiense de Europa central y occidental (Goyet en Bélgica, n = 6 individuos) es tanto geográfica como genéticamente intermedia entre los grupos de Věstonice y Fournol.
LGM en el suroeste y oeste de Europa
La cultura solutrense es intermedia entre las culturas gravetiense y magdaleniense, y se encuentra en el suroeste y oeste de Europa, considerados refugios climáticos para poblaciones humanas durante el LGM.
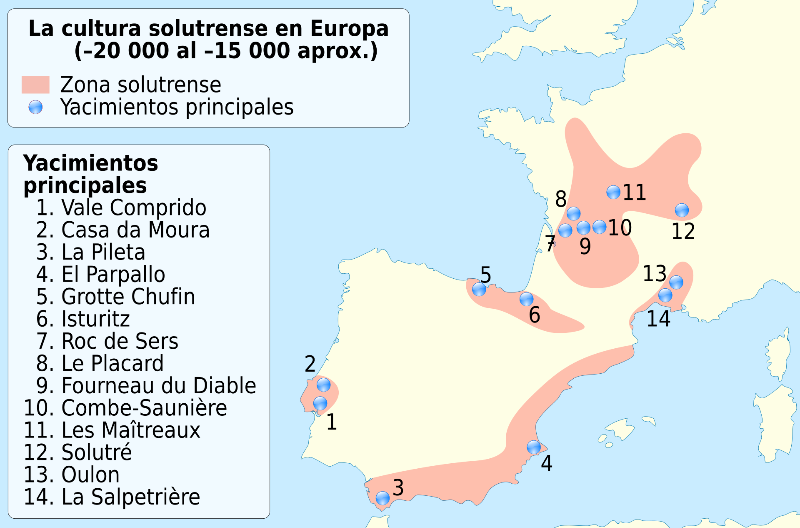
Se desconoce hasta qué punto los grupos del solutrense están en continuidad genética con poblaciones anteriores y posteriores de la misma región, porque no se han estudiado previamente datos genómicos de individuos asociados al solutrense.
Recientemente tienen genomas secuenciados de individuos del solutrense (Le Piage II (23 ka) del suroeste de Francia y La Riera en Posada de Llanes-Asturias (nivel 14, 21 ka) muestran una gran afinidad con miembros Fournol y GoyetQ2.
Le Piage II se sitúa especialmente cerca de los individuos que pertenecen al grupo Fournol, lo que sugiere una continuidad genética local de esta ascendencia durante el LGM. Está más estrechamente relacionado con el grupo Fournol que con el grupo Vestonice. Lo compararon con El Mirón (Cantabria), el individuo asociado al Magdaleniense más antiguo secuenciado hasta la fecha (19 ka) y las estadísticas sugieren que Le Piage II es genéticamente intermedio entre Fournol85 y El Mirón.
Estudios previos habían demostrado que El Mirón llevaba una contribución genética del grupo Villabruna, que se encuentra en individuos asociados al Epigravetiense de Italia. El Mirón tiene una similitud significativamente mayor con el grupo Villabruna que Fournol 85 y Le Piage II.
Este Le Piage II asociado al Solutrense vincula la ascendencia precedente de Fournol con la ascendencia sucesiva encontrada en El Mirón, lo que proporciona evidencia directa de la continuidad genética a lo largo del LGM en el suroeste y oeste de Europa. Por lo tanto, estas regiones europeas constituyen refugios climáticos donde las poblaciones humanas sobrevivieron durante el LGM.
Post-LGM en la península itálica
Después del LGM, la cultura Epigravetiense se extendió por el sur y sureste de Europa.
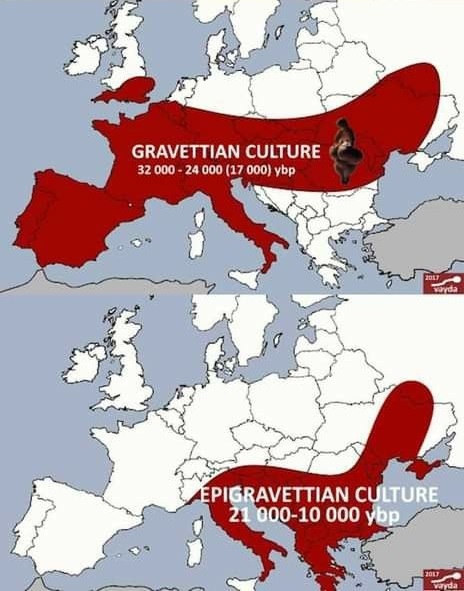
Aquí, nos informan de datos genómicos de 4 individuos nuevos, incluidos 3 genomas de aproximadamente 13.000 años de antigüedad del noreste de Italia (Pradis 1), el noroeste de Italia (Arene Candide 16) y Sicilia (San Teodoro 2), así como una mayor cobertura del genoma completo desde Tagliente 2 datado en 17 ka.
Todos los individuos asociados al Epigravetiense, tanto nuevos como previamente informados, se encuentran dentro del grupo de Villabruna. Estos resultados destacan un recambio genético en la península italiana del grupo Vestonice, asociado al Gravetiense, por el grupo Villabruna, asociado al Epigravetiense, que podría correlacionarse con las discontinuidades observadas en el registro arqueológico. Demostraron que todos los individuos asociados al Epigravetiense analizados tienen una ascendencia Villabruna homogénea. La reconstrucción filogenética de los genomas asociados al Epigravetiense indica que el recambio tuvo lugar mucho antes de 17 ka. Esto, y la evidencia de ascendencia Villabruna en El Mirón 19ka, sugiere además que esta discontinuidad genética podría ser el resultado de transformaciones paleogeográficas y paleo-ecológicas conectadas con el LGM.
La afinidad genética del grupo Villabruna con las ascendencias antiguas y actuales del Cercano Oriente sugiere a los Balcanes como una fuente de la población entrante asociada al Epigravetiense. Puede que el LGM podría haber creado un corredor al sur de los Alpes para los movimientos humanos de este a oeste que conectaron genéticamente a las poblaciones de cazadores-recolectores de los Balcanes con Iberia.
Post-LGM en Europa occidental y central
Los estudios previos identificaron dos mezclas genéticas diferentes en individuos asociados al Magdaleniense:
- el grupo GoyetQ2 que incluye genomas de Europa central y occidental que datan de alrededor de 15 ka (de Francia, Bélgica y Alemania)
- el grupo de El Mirón de España de alrededor de 19 ka.
Ambas ascendencias tienen un componente genético lejanamente relacionado con el individuo Goyet Q116-1 que data de 35 ka, y el individuo ibérico también muestra afinidad con el grupo Villabruna .
Al analizar datos publicados previamente con nuevos genomas asociados con el Magdaleniense (La Marche , Pincevent y Maszycka ), confirmaron que la ascendencia Goyet Q116-1 sobrevivió en todos los genomas asociados al Magdaleniense estudiados además de en individuos asociados al Gravetiense y Solutrense del suroeste y oeste de Europa. Y demostraron que todos los individuos asociados al Magdaleniense, y no solo El Mirón, tienen ascendencia relacionada con Villabruna. El Mirón presenta alrededor de un 43% de ascendencia de Villabruna.
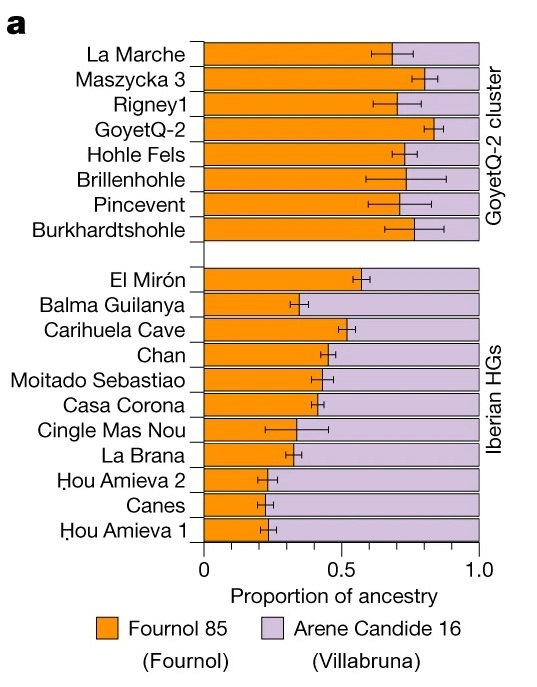
Imagen: a , b , ascendencias de los individuos del grupo GoyetQ-2 y de los cazadores-recolectores (HG) ibéricos ( a ) y de los individuos del grupo Oberkassel ( b ) se modelaron con Fournol 85 y Arene Candide 16 representando las ascendencias de Fournol y Villabruna respectivamente.
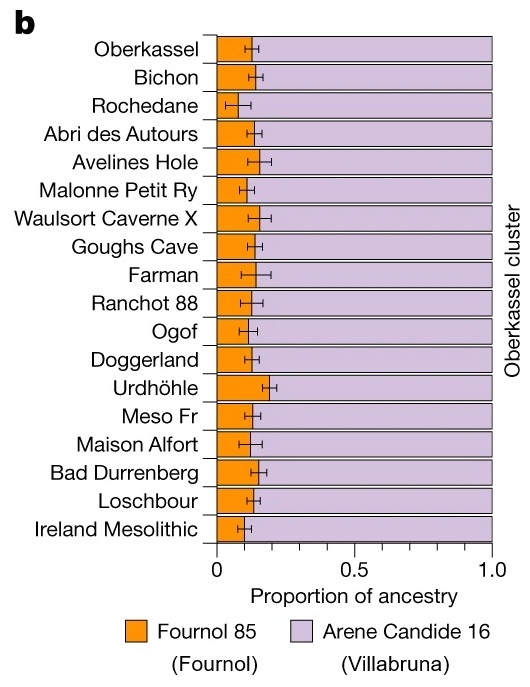
Estos análisis demuestran que el grupo Fournol es una mejor fuente de genomas asociados al Magdaleniense que Goyet Q116-1. Por lo tanto, la mayor parte de la ascendencia encontrada en estos individuos post-LGM probablemente se remonta a grupos asociados al Gravetiense de Europa occidental y sudoccidental.
La afinidad genética con la ascendencia de Villabruna está presente en El Mirón y en individuos asociados al Magdaleniense de Europa occidental y central.
Esto sugiere que los cazadores-recolectores del sur y suroeste de Europa en la época del LGM se extendieron al norte de los Pirineos. El grupo GoyetQ2 resultante incluye individuos que abarcan desde el oeste de Francia hasta Polonia en el período entre 18 y 15 ka. Por lo tanto, esto demuestra que la difusión post-LGM del Magdaleniense estuvo de hecho asociada con expansiones poblacionales hacia el norte y el noreste desde Europa occidental.
Post-14 ka al Neolítico
Estudios previos habían demostrado que dos grupos principales de cazadores-recolectores fueron predominantes en la mayor parte de Europa después de alrededor de 14 ka, es decir, el grupo occidental de cazadores-recolectores (WHG) relacionado con el grupo Villabruna, y el grupo oriental de cazadores-recolectores (EHG), que muestra afinidad tanto con Villabruna como con el ancestro antiguo del norte de Eurasia (ANE) encontrado en individuos siberianos del Paleolítico Superior .
La mayoría de los individuos post-14 ka de Europa occidental y central caen cerca del grupo WHG y aquellos de Europa oriental cerca del grupo EHG.
Los dos individuos de Oberkassel de 14 ka marcan la presencia más temprana de ascendencia WHG al norte de los Alpes. Estos individuos Oberkassel están más cerca del genoma de Arene Candide 16 (Villabruna) que cualquier otro grupo asociado al Epigravetiense de Italia. Además, el grupo de Oberkassel lleva tanto la ascendencia Villabruna como una contribución de la ascendencia GoyetQ2.
La observación de que los individuos posteriores a 14 ka de Europa occidental y central y de Gran Bretaña llevan una composición genética homogénea implica que el perfil de ascendencia Oberkassel ya estaba formado en gran medida antes de su dispersión. Esto contrasta con la historia genética de los cazadores-recolectores ibéricos, donde la expansión de la ascendencia Villabruna/Oberkassel implicó múltiples mezclas locales con grupos que portaban altas proporciones de ascendencia GoyetQ2.
La prolongada continuidad genética en Iberia se refleja en la preservación hasta el Mesolítico del haplogrupo C del cromosoma Y, que fue predominante en los grupos pre-LGM, pero rara vez se encontró después del LGM en otras partes de Europa.
Confirmaron que las poblaciones EHG en Europa del Este son una mezcla de ancestros Villabruna/Oberkassel y ANE. Este estudio muestra que el grupo Yuzhniy Oleniy Ostrov de Karelia en Rusia occidental de aproximadamente 8,2 ka formado por 19 genomas tiene una afinidad comparable o menor con el ancestro Villabruna que todos los demás grupos EHG.
Luego modelaron 250 cazadores-recolectores publicados y los recientemente encontrados fechados en 14-5 ka usando mezclas de ascendencias Oberkassel, Sidelkino, GoyetQ2 y una ascendencia de agricultores neolíticos de Anatolia (ANF), ya que una porción considerable de los genomas de cazadores-recolectores secuenciados datan de alrededor de 8 ka, cuando la ascendencia ANF comenzó a extenderse por Europa.
Estos resultados muestran que la zona de contacto y algunos patrones de mezcla entre las ascendencias de Oberkassel y Sidelkino cambiaron con el tiempo.
Entre 14 y 8 ka, todos los cazadores-recolectores en Europa occidental y central portaban solo ascendencia de Oberkassel, sin contribución detectable del grupo Sidelkino.
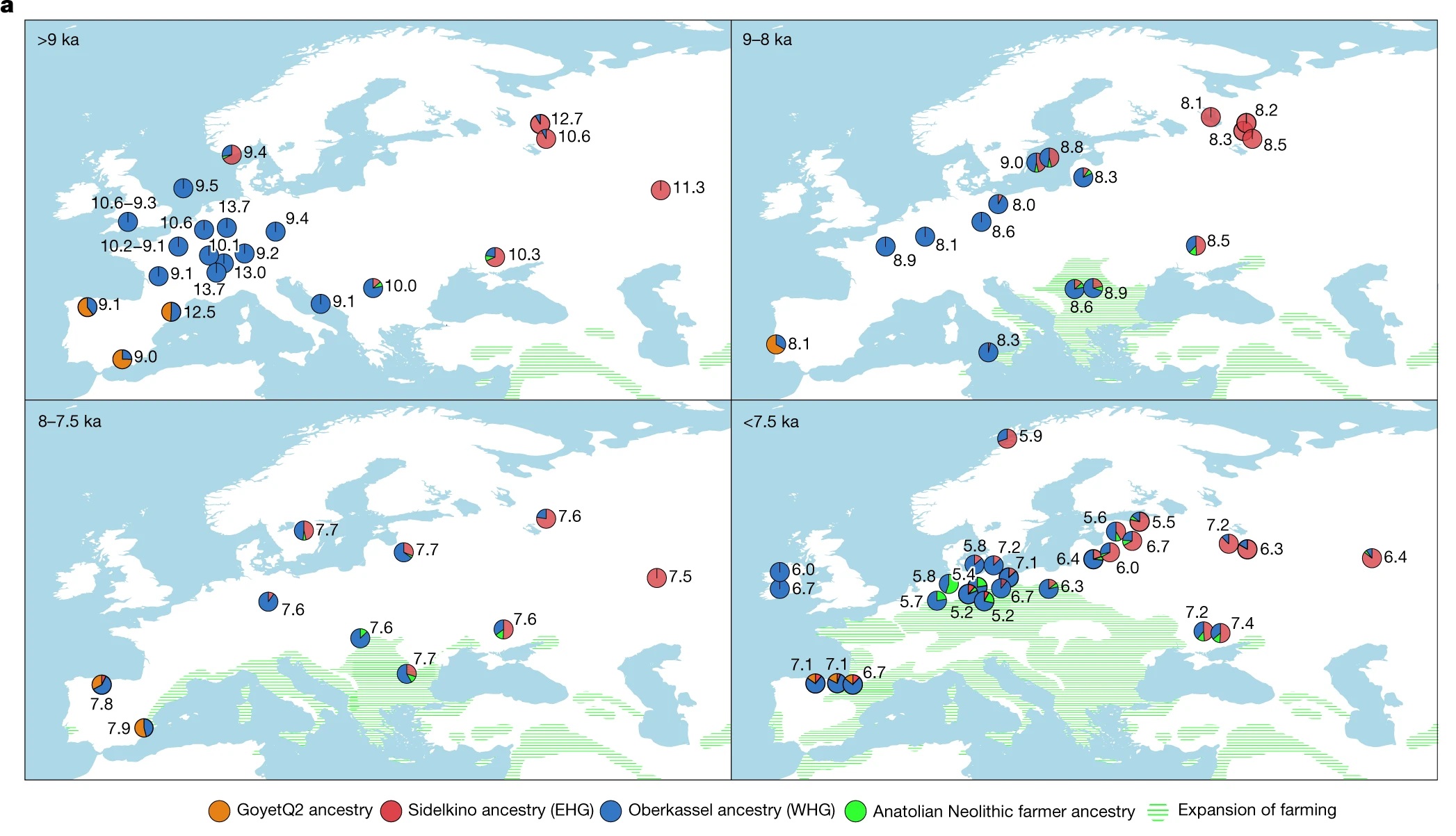
Imagen a , La ascendencia genética de los cazadores-recolectores datada entre 14 ka y 5,2 ka
modelada utilizando qpAdm, con Oberkassel, Yuzhniy Oleniy Ostrov, Goyet Q-2 y agricultores
neolíticos de la actual Turquía (sitios de Barcın, Menteşe y Boncuklu) representando las
ascendencias de Oberkassel (WHG) (azul), Sidelkino (EHG) (rojo), GoyetQ2 (naranja) y
agricultores neolíticos de Anatolia (verde), respectivamente. Se muestra la fecha calibrada
promedio, con gráficos circulares que indican la proporción estimada de ascendencia para cada
grupo o individuo. La expansión de la agricultura en 9, 8, 7,5 y 7 ka se muestra como tonos verdes. Adaptado
de https://doi.org/10.5281/zenodo.5903165 (CC BY 4.0).
Dos de los grupos publicados más antiguos de Rusia occidental pertenecientes al grupo Sidelkino, Peschanitsa (13 ka) y los individuos Minino recientemente publicados (11 ka), mostraron una afinidad extra con el grupo Oberkassel. Estimaron la mezcla entre las ascendencias de Villabruna/Oberkassel y ANE en estos antiguos individuos relacionados con el grupo Sidelkino en alrededor de 15-13 ka. Coincide con la aparición de la ascendencia de Oberkassel en Europa central y esto plantea la posibilidad de que la sustitución por el grupo Oberkassel y la formación del grupo Sidelkino pudieran haber sido el resultado de expansiones poblacionales influenciadas por el calentamiento abrupto durante el interestadial Bølling-Allerød .
A partir de alrededor de 8 ka, comenzamos a observar eventos de mezcla con ascendencia Sidelkino en Europa central, que alcanza alrededor del 10% en la mayoría de los individuos cazadores-recolectores europeos a partir de entonces.
Poco después de 8 ka, la ascendencia Sidelkino estaba ausente en el este de España, pero ya había llegado al norte de Iberia junto con un aumento en la ascendencia Oberkassel. Por el contrario, se identifica ascendencia Oberkassel adicional en Europa oriental por al menos 7,5 ka en genomas recién generados de Minino I y Yazykovo de la región superior del Volga.
Después de 7,5 ka, la ascendencia Anatolia ANF había alcanzado regiones al norte de los Alpes, los individuos que llevaban un perfil genético de cazadores-recolectores se restringieron principalmente a las franjas septentrionales de Europa.
En este período, la mezcla de ascendencia Oberkassel se extendió más al este, llegando a Samara alrededor de 6,5 ka, y se detectó un aumento en la ascendencia Sidelkino en cazadores-recolectores de la región del Báltico, que anteriormente se asoció con la transición de la cultura Narva a la cultura de cerámica de peine.
En Europa central, la mezcla con ascendencia ANF se volvió muy común pero no ubicua, lo que indica la coexistencia de sociedades de cazadores-recolectores y agricultores sin mezclarse durante varios cientos de años. El individuo más joven con gran proporción de ascendencia cazadora-recolectora en el conjunto de datos analizado procede de Ostorf, en el norte de Alemania, y data de alrededor de 5,2 ka (más del 90 % del cúmulo de Oberkassel y componentes del cúmulo de Sidelkino). Los individuos de este yacimiento podrían marcar una de las últimas apariciones de niveles tan altos de ascendencia relacionada con cazadores-recolectores, apenas siglos antes del inicio de la Edad del Bronce europea.
Un individuo neolítico Tutkaul 1 de Tayikistán está estrechamente relacionado con los individuos del Paleolítico superior del centro sur de Siberia (Afontova Gora 3 (AG3) y Mal'ta 1), y con cazadores-recolectores siberianos occidentales. Probaron la afinidad de Tutkaul 1 con las poblaciones antiguas y modernas de todo el mundo en relación con AG3. A diferencia de los cazadores-recolectores siberianos occidentales, Tutkaul 1 no tiene una ascendencia euroasiática extra oriental, pero muestra afinidad con los agricultores neolíticos iraníes y algunas poblaciones más jóvenes de Irán y la región de Turan.
Variantes fenotípicamente relevantes
En consonancia con hallazgos previos, ninguno de los grupos analizados muestra el alelo derivado en el SNP rs4988235 del gen LCT responsable de la persistencia de la lactasa.
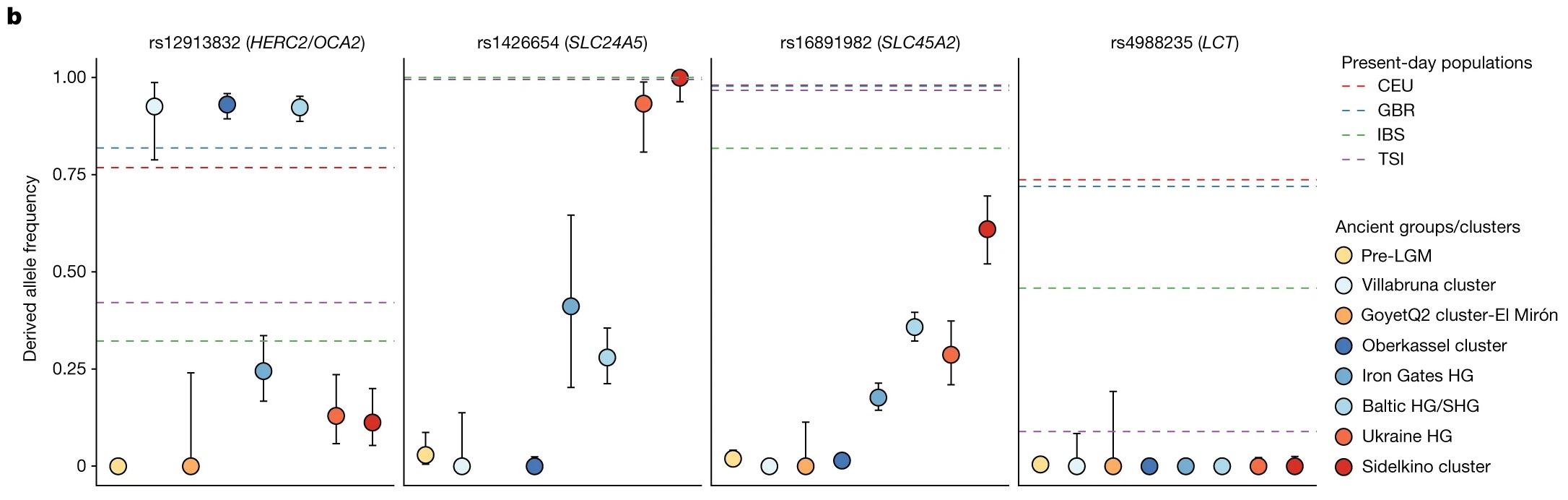
b Frecuencias alélicas de diferentes grupos de cazadores-recolectores (puntos de colores) en cuatro SNP relacionados con el color de la piel ( SLC24A5 y SLC45A2 ), el color de los ojos ( HERC2 / OCA2 ) y la persistencia de la lactasa ( LCT ). Los puntos son estimaciones de máxima verosimilitud y las barras de error muestran intervalos de confianza del 95% de las frecuencias alélicas derivadas. Las líneas discontinuas muestran las frecuencias estimadas para las poblaciones actuales indicadas del Proyecto 1000 Genomas (CEU, residentes de Utah de ascendencia europea del norte y oeste; GBR, británicos; IBS, españoles; TSI, toscanos ) .
El SNP asociado con el color de ojos claros ( HERC2/OCA2 (rs12913832)), los individuos del grupo Villabruna, el grupo Oberkassel, los grupos Baltic HG y SHG muestran frecuencias altas del alelo derivado (>90%), que es responsable del fenotipo de ojos verdes o azules, mientras que los grupos Sidelkino, Ukraine HG y Iron Gates HG muestran una baja incidencia de este alelo (10-25%).
Los dos SNP asociados con el color de la piel ( SLC24A5 (rs1426654) y SLC45A2 (rs16891982)), aparecen en el grupo Sidelkino y los grupos Ukraine HG en una frecuencia más alta (>90% para SLC24A5 y 29-61% para SLC45A2 ), en comparación, los grupos Oberkassel y Villabruna esos alelos están casi completamente ausentes (<1%).
Discusión y conclusiones
La cultura Gravetiense en toda Europa no era una población biológicamente homogénea. La ascendencia de individuos asociados con la cultura Auriñaciense precedente de Europa central (ascendencia GoyetQ116-1) dio lugar a individuos asociados con el Gravetiense de Europa occidental y suroccidental. Esta ascendencia derivada, el grupo Fournol, sobrevivió durante el LGM en individuos asociados al Solutrense, posiblemente dentro del refugio climático franco-cantábrico , lo que dio lugar a poblaciones posteriores asociadas con la cultura Magdaleniense (grupo GoyetQ2 y El Mirón).
La ascendencia de los individuos de la cultura Epigravetiense (grupo Villabruna), que se descubrió que conectaba genéticamente a los cazadores-recolectores europeos y de Oriente Próximo, llegó al sur de Europa mucho antes de la transición entre el Epigravetiense Temprano y el Tardío y posiblemente incluso en la transición Gravetiense-Epigravetiense.
Los individuos asociados al Magdaleniense, no solo de Iberia, sino también del resto de Europa, poseen ascendencia asociada al Epigravetiense (grupo Villabruna). Los análisis genéticos de individuos de Europa occidental asociados con la cultura Badegouliense precedente podrían proporcionar pistas sobre los procesos que llevaron a la formación del grupo GoyetQ2.
Como se infiere del registro arqueológico, la expansión del Magdaleniense por Europa está vinculada a las expansiones poblacionales del suroeste al norte y noreste posteriores al LGM, y no a movimientos desde refugios del sureste.
Este estudio revela que Europa occidental y suroccidental sirvieron como refugios climáticos para la persistencia de grupos humanos durante la fase más fría de la última Edad de Hielo, mientras que las poblaciones de la península itálica y la llanura europea oriental sufrieron un cambio genético, lo que pone en entredicho el papel de estas regiones como refugios glaciares para los humanos.
La ascendencia Villabruna, que llegó a Europa, se convirtió posteriormente en la ascendencia de cazadores-recolectores más extendida en toda Europa.
Estudios paleogenómicos adicionales sobre individuos del Paleolítico Superior de los Balcanes serán esenciales para comprender si el sureste de Europa representa el origen de la ascendencia Villabruna y un refugio climático para las poblaciones humanas durante el UGM.